Titelbild : Kormoraneinflug an der Nister 19.10.2019 um 8:18 Uhr
Nach mündlicher Rücksprache und freundlicher Genehmigung von Martin Görner dürfen wir diesen Artikel auf unsere Homepage setzen.
Spannungsfeld zwischen Kormoran (Phalacrocorax carbo) und Ziel-stellungen des Artenschutzes
Martin Görner
Abstract: The increasing population of cormorants in Europe since 1990 caused conflicts between the aims of bird, fish
and biotope conservation issues.
The focus of this paper is on conservation of biodiversity. The aim is to preserve the inland populations of both cormorant
and fish. The fish species of the biotopes trout-, grayling- and barb-region are particularly affected.
Insights need to be attained to resolve species conservation problems.
Key words: Cormorant, Phalacrocorax carbo, population dynamics, predator, ecology, habitat, fishes, conservation
1. Einleitung
Spätestens seit 1980 hat der Kormoran (Phalacrocorax carbo sinensis) in seinem Brutbestand
europaweit erheblich zugenommen. Zu dieser Zeit haben die Vögel ihre Brutgebiete, und dies war neu, besonders im Binnenland, wie beispielsweise in Deutschland, ausgedehnt. Der Kormoran jagt von den Meeresküsten Europas bis nunmehr in Bächen, Flüssen und Seen des tiefen Binnenlandes, auch das ganze Jahr über. Mit der nachgewiesenen Abnahme von Fischen durch Kormorane in diesen Gewässern sowie in Fischzuchtanlagen entstehen Konflikte, die je nach Standpunkt der Betrachter unterschiedlich bewertet werden.
Die oft gestellte Frage, wie groß der Fraßdruck des Kormorans auf den Fischbestand in Bächen
und Flüssen, aber auch in Fischzuchtanlagen, ist, kann in der Mehrzahl der Fälle durch seriöse
wissenschaftliche Untersuchungen von Fischereibiologen beantwortet werden. Der enorme Populationsanstieg des Kormorans in den letzten Jahrzehnten beeinflusst die Fischbestände in Fließgewässern teilweise erheblich. Hierzu liegen ausreichende Gutachten und Veröffentlichungen aus verschiedenen Gebieten Mitteleuropas vor (z. B. Bokranz et al. 1998, Brämick 2005, Bzoma et al. 2005, Conrad et al. 2002, Dauste r 1987, Dornbusch & Fische r 2010, Görlach & Wagner 2006, Jungwirth 1995, Kainz 1994, Keller 1997, Keller et al. 1996, Kieckbusch 1993, Klein 2000, Kohl 2005, Müller 1986, Neubauer et al. 2017, Ruh le 1985, Schmalz & Schmalz 2003, Schwevers & Adam 1998, Staub 1992, Wagner et al. 2008, Worthma nn & Spratte 1990).
Es muss nicht noch weiter geforscht werden, um den Einfluss des Kormorans auf die Fischfauna zu belegen. Offensichtlich fällt eine Objektivierung der Problemwahrnehmung bei Naturschützern, Ornithologen, Fischern und Anglern, aber auch Behördenmitarbeitern schwer. Reichen die möglichen und viel diskutierten Präventionen aus, um die anstehenden Probleme im Artenschutz einer Lösung näher zu bringen? Der Zielsetzung, die Biodiversität zu erhalten und nach Möglichkeit zu fördern, stimmen die unterschiedlichen Betrachter zu, bieten aber kaum eine tragbare und akzeptable Lösung an. Es muss nicht besonders hervorgehoben werden, dass wohl alle Fischbestände einem mehr oder weniger starken Fraßdruck durch fischfressende Prädatoren unterliegen. Bless (1989) hat darauf hingewiesen, dass die Schutzproblematik der Fische und Rundmäuler noch bis in die neueste Zeit sowohl in der Naturschutzgesetzgebung als auch in der praktischen Arbeit vernachlässigt wurde. Zu dieser Zeit war aber die Zunahme der Kormoranpopulationen mit ihren Problemen schon in der öffentlichen Diskussion.
Acta ornithoecologica, Bd. 9, H. 1, 2019
Acta ornithoecologica, Jena 9. 1 (2019) 59 – 87 30.03.2019 59
Ornithologen und Naturschützer bemühen sich um den Schutz der Vögel. Fischereibiologen,
Fischer und Angler bemühen sich um den Schutz der Fische. Artenschützer, Wildbiologen und
Ökologen haben die Kormorane und die betroffenen Fischarten auf gleicher Ebene in den Blick
zu nehmen, da es um die Erhaltung der „Biologischen Vielfalt“ geht. Warum fallen der sachliche Austausch und die Anerkennung von wissenschaftlichen Fakten – es geht nur um diese – der Mehrzahl der jeweiligen Betrachter so schwer? Wenn der Bestand einer Art rapide abnimmt (z. B. Äsche) und die Ursachen sichtbar und eindeutig belegt sind (z. B. verstärkte Prädation), muss gegengesteuert werden, wenn die Zielstellung „Erhaltung der Biologischen Vielfalt“ ernst gemeint wird. Diesem Prinzip sollte sich auch die Politik nicht verschließen, selbst wenn einzelne Maßnahmen zunächst nicht populär sind. Die öffentliche Meinung ist in Sachen Kormoran und vieler weiterer Arten derart angeheizt, dass offensichtlich fachliche Argumente nicht mehr aufgenommen, überdacht und verarbeitet werden. Die Ziele des Naturschutzes ergeben sich aus den Bestimmungen des Bundesnaturschutzgesetzes sowie den verschiedenen europäischen Richtlinien, wobei besonders der Schutz der Arten und somit die Biologische Vielfalt (Biodiversität) im Mittelpunkt stehen müssen. Die Betrachtungen gelten allen Arten, unabhängig von Lebensweise, Körpergröße, Attraktivität, vom Bekanntheitsgrad, von Häufigkeit und Wahrnehmbarkeit.
2. Zur Entwicklung des Kormoranbestandes in Deutschland
Bereits Nauma nn (1817) schreibt u. a.: „Er [der Kormoran] kommt sowohl an der Ostsee wie
auch an der Nordsee vor und ist in Holland ziemlich häufig, aber auf die Gewässer des inneren
Deutschlands kommt er nur höchst selten, und mehrentheils nur in den Wintermonaten.“
Hennicke (1912) führt zur Verfolgung von Vögeln aus wirtschaftlichen Gründen u. a. aus,
dass der Kormoran als Schädiger der deutschen Binnenfischerei schon deshalb nicht anzusehen
ist, weil „bei uns in Deutschland nur eine einzige Kolonie bekannt ist und die paar im Winter ins
Binnenland verirrten Vögel wirtschaftlich überhaupt nicht in Betracht kommen können.“
Nach Niethamme r (1940) war der Kormoran (Phalacrocorax carbo) in allen seenreichen
Gebieten Norddeutschlands von Ostpreußen bis Nordschleswig ein Brutvogel: „Westlich der
Elbe und in Süddeutschland fehlen die fischreichen Gewässer in genügender Anzahl und Ausdehnung; soweit bekannt, haben Kormorane dort nie gehorstet.“ Bauer & Glutz von Blotzheim (1966) haben für Mitteleuropa ab dem 18. Jahrhundert bis 1965 Brutplätze und -bestände des Kormorans dargestellt. Die auf dem heutigen deutschen Staatsgebiet liegenden alten bzw. ehemaligen Brutplätze sind aus Abb. 2 ersichtlich. Seit dem 16. Jahrhundert sind meist einzelne Brutvorkommen des Kormorans in den Grenzen der aktuellen Bundesländer Schleswig-Holstein, Mecklenburg-Vorpommern, Brandenburg, Niedersachsen, Sachsen-Anhalt und Sachsen belegt. Weitere Einzelheiten dazu vgl. z. B. Herrmann (2011 u. 2012). In diesem Zusammenhang sei auch hervorgehoben, dass archäologische Funde vom Kormoran im europäischen Binnenland bekannt wurden (vgl. Heinrich 2007, Kinzelbach 2010). Beike (2012) konnte nachweisen, dass der Kormoran in einzelnen Ländern Europas im 16. und 17. Jahrhundert vom Menschen zum Fischfang eingesetzt wurde.
Jahrzehntelang hat die menschliche Verfolgung der Kormorane in verschiedenen deutschen
Brutgebieten einen erheblichen Einfluss auf den Brutbestand gehabt. Von den wenigen Kolonien
im heutigen Mecklenburg-Vorpommern hat die Art etwa ab 1950 infolge strengen Schutzes wieder zugenommen. Die Vögel brüteten lokal konzentriert an der Küste und an den Binnengewässern des Landes. 1963 waren über 1.000 und 1983 etwa 1.300 Brutpaare bekannt (vgl. Zimmermann 1987). Mit der teilweisen Einstellung der Verfolgung von Vögeln in den Brutgebieten und dem Rückgang der Pestizidbelastung in der Umwelt nach 1970 nahm der Kormoranbestand, auch durch die 60 Acta ornithoecologica, Bd. 9, H. 1, 2019 EG-Richtlinie 79/409 EWG über die wildlebenden Vogelarten vom 02.04.1979, in ganz Europa wieder zu. In der genannten Richtlinie wird der Kormoran in den Anhang I eingeordnet. Somit oblag allen EU-Ländern die Verpflichtung, die Art ganzjährig zu schützen (bezieht sich auf die Brut-, Rast- und Überwinterungsgebiete). Dies löste einen europaweiten deutlichen Anstieg aus, was als ein Erfolg des Naturschutzes gewertet werden darf, da besonders durch rechtliche Schutzmaßnahmen (Abschussverbot, Beunruhigung am Brutplatz) die größte Gefahr von den Vögeln genommen wurde. Die Kormoranbestände waren bis Anfang der 1970er Jahre nahezu ausschließlich an den Meeresküsten und den Flussmündungen Mitteleuropas konzentriert. Die im Binnenland an den unterschiedlichen Gewässern vorherrschenden oft durch Eutrophierung ausgelösten und für die Vögel häufig günstigen Nahrungsbedingungen hatten ebenfalls einen positiven Einfluss auf den Kormoranbestand.
Mit der flächenhaften Ausweitung der bisherigen Vorkommensgebiete im deutschen und mitteleuropäischen Binnenland wurde auch die Flexibilität der Vögel hinsichtlich der Brutplätze
(z. B. Bodenbrüter auf Inseln und Baumbrüter) deutlich sichtbar. Die ehemals klassischen Brutbereiche des Kormorans in Deutschland wurden in Tab. 1 dargestellt (vgl. Bundesländer Nr. 1 bis 5). Abb. 1 Vereinfachte Nahrungspyramide von fischfressenden Vögeln und Säugetieren im mitteleuropäischen Binnenland. Kormorane fangen Fische; von der zwischen Steinen lebenden, kleinen, bodenbewohnenden Groppe (Cottus gobio) bis zu etwa 45 cm langen Barben (Barbus barbus) (Entwurf: M. Görner, Grafik: K. Maltzah n). Acta ornithoecologica, Bd. 9, H. 1, 2019 61
Abb. 2 Räumliche Verteilung der bekannten und wieder aufgegebenen Kormoranbrutplätze im Küstenbereich Deutschlands und dessen Hinterland bis etwa 1970: nummerierte rote Punkte siehe Tab. 1 (vgl. auch BAUER & GLUTZ von BLOTZHEIM 1966), orangefarbige Punkte: ehemalige Brutplätze, die nach 1950 bestanden (vgl. Herrma nn 2012). 62 Acta ornithoecologica, Bd. 9, H. 1, 2019 Tab. 1 Nachweise von Bruten, Zugbewegungen und Winterbeständen des Kormorans in Deutschland nach den heutigen Bundesländern (alte Nachweise vgl. Abb. 2).
Acta ornithoecologica, Bd. 9, H. 1, 2019 63 Nr. Bundesland Nr. Ortsbezeichnung Zeit(raum) Bemerkung 1 Schleswig-Holstein 1 Schlei bei Kappeln Nach 1816 Nach Vernichtung der Kolonie
der Hohwachter Bucht Umzug in Reiherkolonie an der Schlei, Vertreibung und Unterbindung weiterer Brutversuche
2 Buckhagen / Schlei 1943 und 1944 Je 3-4 Paare in Fischreiherkolonie;
Verschwinden nach 2 Wochen
3 bei Itzehoe Noch 1881 In Reiherkolonie Julianka; nur kurze
Dauer, da gleich unterbunden
4 Binnensee der Hohwachter
Bucht
1810-1816 In Saatkrähen- bzw. Reiherkolonie;
1816 Zerstörung der Kolonie angeordnet und ausgeführt (Vernichtung von 1.660 Nestern)
Ehemals Brutvogel, gab es um 1974 (Berndt & Drenckhah n) bis 100
„Durchzügler“ (Wintervögel 10-50 Ex.). Die Autoren führen aus: „Trotz der umliegenden Kolonien und Rastplätze ist der Durchzug des Kormorans in Schleswig-Holstein relativ gering“.
Im Jahr 2016 waren 2.254 BP bekannt (Deutscher Bundestag 2017).
2 Mecklenburg- Vorpommern
5 Hohen Wischendorf an der Ostsee
Vor 1900 Zeitweise sehr große Kolonie
6 Pinnower See bei Schwerin
Vor 1900 Zeitweise sehr große Kolonie
7 Bolzer See (nordöstlich
von Borkow)
1965 Etwa 25 Paare
8 Fischland
Vor 1900und 1922
Zeitweise sehr große Kolonie;
1922: einige Paare in Fischreiherkolonie
9 Moorsee bei Waren / Müritz
1912 und 1913 Brut von 6 bis 7 Paaren
10 Müritzsee
Vor 1900 Zeitweise sehr große Kolonie
11 Tollensesee
Vor 1900 Zeitweise sehr große Kolonie
12 Niederhof bei Brandshagen am Strelasund
1954 Kolonie
1954 entdeckt: ca. 60
Vögel (wohl Auswanderer von Rügen); in Folgejahren vergrößert
13 Insel Altrügen 1946(-1950?) Übersiedlung von Insel Pulitz,
aber keine erfolgreiche Brut;
1947: 200-250 Paare,
1948: ca. 400 Paare (trotz Störungen),
1949: 200 Nester (nur 10 erhalten);
1950: letzte Brut auf Rügen, 500 Vögel, aber nur Brut von 10-12 Paaren
Tab. 1 (Fortsetzung)
64 Acta ornithoecologica, Bd. 9, H. 1, 2019
Nr. Bundesland Nr. Ortsbezeichnung Zeit(raum) Bemerkung
14 Insel Pulitz
(Jasmunder
Bodden)
Mind. ab 1939
bis 1945
1939: 350-400 BP, in Folgejahren
500-600 BP, 1945: Verhinderung
der Brut durch Störungen
15 Usedom 1938 Neuansiedlung; 1939: 20 Paare
Mehrere ältere Quellen weisen auf die Brutplätze des Kormorans und die
Bestandsentwicklung in diesem heutigen Bundesland hin (vgl. z. B. Home yer
1837, Maltzan 1848, Zande r 1862, Kuhk 1939, Baue r & Glutz von
Blotzheim 1966). In diesem Zusammenhang sei auch besonders auf die umfassende
Zusammenstellung von historischen Angaben für Mecklenburg und Pommern
vom ausgehenden 18. bis zur Mitte des 20. Jahrhunderts hingewiesen (vgl.
Herrma nn 2011). Weitere informative Details zur Situation des Kormorans,
die auch das o. g. Bundesland betreffen, hat Herrma nn (2012) veröffentlicht.
Im Jahr 1980 betrug der Brutbestand ca. 705 (Herrma nn 2012), 2005 ca.
12.765 (Heinicke 2005), 2008 ca. 14.400 und 2016 ca. 15.400 Paare (Deutscher
Bundestag 2017).
3 Brandenburg 16 Bei Fürstenberg Vor 1900 Zeitweise sehr große Kolonie
17 Werder 1965 Brut von 6 Paaren am Havelufer
Nach Scha low (1919) dürfte sich der Kormoran erst ungefähr seit dem ersten
Jahrzehnt des 19. Jahrhunderts als nicht gerade seltener Brutvogel in Brandenburg
angesiedelt haben: „Am Werbelinsee bei Joachimsthal ließ sich 1837 eine ungeheure
Kolonie nieder“. Schließlich wurde das Garde-Jäger-Bataillon aus Potsdam
eingesetzt, um die Vögel zu vertreiben.
Interessant ist der Hinweis bei Scha low (1919), „[…] dass von der Oderburger
Kolonie der Aal in sämtlichen Seen der Umgegend fast völlig ausgerottet worden
sei; dasselbe gilt von den Maränen (Coregonus generosus) des Werbelinsees“.
Nach Rutschke (1983) war der Kormoran als Brutvogel im 19. Jahrhundert
gemeinsam mit Graureihern in Kolonien bekannt. 1965 siedeln sich erneut Kormorane
an; erste Brutversuche wieder 1971. 1977 gab es Nachweise an den Peitzer Teichen.
1977 siedelten mehrere Brutpaare in einer Reiherkolonie im Kreis Hoyerswerda.
Im Jahr 2010 gab es nach Ryslav y (2011) in Brandenburg 2.600 Brutpaare.
2016 waren es ca. 1.400 BP (Deutscher Bundestag 2017).
4 Niedersachsen 18 Lütetsburg / Kreis
Norden (Ostfriesland)
Seit 1944, bis
etwas nach
1958
In Fischreiherkolonie; 1944: 1 BP,
1945: 3 BP, 1946: 7 BP, 1947: 15 BP,
1951: 59 BP, 1952: 70 BP, 1957:
83 BP; 1958: 40 BP (Reduzierung
durch Abschuss), später erloschen
19 Eversand Seit mindestens
1947
Auf zwei alten Leuchttürmen;
1957: 20 besetzte Nester,
1959: etwa 50 Paare
20 Meyers Legde
(Wesermündungsgebiet)
Seit mindestens
1947
Auf Leuchtturm; im Frühjahr 1960:
46 Paare
21 Vor der Wesermündung
1924 Nisten von 3 Paaren
Die Situation des Kormorans in den anderen Bundesländern kann wie folgt skizziert werden:
6 Nordrhein-Westfalen
Kormoranbeobachtungen gibt es aus dem gesamten Rheinland vor der Jahrhundertwende. Es
wurden damals Einzelvögel und auch Trupps beobachtet (vgl. Milde nbe rge r 1982).
Nach Neubau r (1957) konnte jedoch ein Brutnachweis nicht erbracht werden, obwohl sich
im Kreis Wesel geschlechtsreife Vögel zur Brutzeit aufhielten. Nach Milde nbe rge r (1982)
nahmen seit Mitte der 1960er Jahre die Beobachtungen, vor allem auf den neu entstandenen und
mit Fischen besetzten Baggerseen, wieder zu.
Für das Niederbergische Land führt Skiba (1993) aus: „Der Kormoran wurde als Durchzügler
und Wintergast bereits im vergangenen Jahrhundert vorwiegend am Rhein, aber auch fernab
davon vereinzelt an geeigneten Gewässern beobachtet. […] Seit 1986 wurden vereinzelte Brutversuche
festgestellt.“
Jöbges & Herke nrath (2017) haben die Vorkommen des Kormorans für dieses Bundesland
von 1986 bis 2016 zusammenfassend dargestellt. Erstmals brütete die Art im Landesteil
Nordrhein im Jahr 1986 und seit 1991 im Landesteil Westfalen (Jöbges 2013). Seit dieser Zeit
vergrößerte sich der Brutbestand bis auf 1.171 Brutpaare im Jahr 2016.
Die Autoren weisen darauf hin, dass der Brutbestand seit 2012 mit 1.100 und 1.200 BP stabil
ist und nicht weiter anwächst.
7 Hessen
Nach Gebha rdt & Sunke l (1954): „treten auch heute noch am Rhein kleinere Gruppen
oder Einzelvögel zu allen Jahreszeiten auf. Ein früheres Brüten der Vögel konnte nicht bestätigt
werden. Die übrigen Teile Hessens werden vom Kormoran nur höchst selten berührt.“
1985 war der Kormoran erstmals Brutvogel. Es erfolgte eine stetige Bestandszunahme bis auf
569 Brutpaare im Jahr 2004, dann von 2010 bis 2015 stagnierte der Bestand mehr oder weniger
um 340 BP (vgl. Brau neis 1999, Cimiotti et al. 2013, Deutscher Bundestag 2017).
Acta ornithoecologica, Bd. 9, H. 1, 2019 65
Nr. Bundesland Nr. Ortsbezeichnung Zeit(raum) Bemerkung
22 Schnackenburg an
der Elbe
2. Hälfte des
19. Jahrhunderts
Wiederholte Brut
Vor 1861 haben bei Schnackenburg Kormorane in einer Reiherkolonie gebrütet
(vgl. Goethe et al. 1978). Die Autoren schätzen ein, dass die Besiedlung der
westdeutschen Brutplätze nach 1945 von den Niederlanden ausgegangen sein
dürfte.
Nach Hecke nroth & Laske (1997) ist der Kormoran mindestens seit 1941
wieder und danach durchgehend Brutvogel. Ab den 1980er Jahren kam es an verschiedenen
Plätzen zur Gründung neuer Kolonien. 2016 wurden 1.330 BP gezählt
(Deutscher Bundestag 2017).
5 Sachsen-
Anhalt
23 Bei Havelberg 1924-1928 Kleine Kolonie; 1925: 6 Paare,
1926: 25 Paare, 1928 erloschen
Seit 1928 brüteten erstmals wieder 1979 5 Brutpaare im Südteil des heutigen Bundeslandes
Sachsen-Anhalt. Im Jahr 2016 betrug der Brutbestand 660 Paare (Deutscher
Bundestag 2017).
Abb. 3 Verteilung von Kormoranbeobachtungen
von 1951 bis 1980 in Thüringen (aus
Schmidt 1986).
Abb. 4 Selbst in Fischaufstiegsanlagen (Weiße Elster in Gera)
fischen Kormorane sehr erfolgreich (Aufn.: S. Heid ler).
Abb. 5 Kormoran beim
Fang einer nahezu ausgewachsenen
Bachforelle
(Aufn.: S. Heid ler).
Abb. 6 Eine stattliche
Barbe wird vom Kormoran
gefangen (Aufn.:
S. Heid ler).
66 Acta ornithoecologica, Bd. 9, H. 1, 2019
8 Thüringen
Vor 1945 gab es insgesamt weniger als 20 Nachweise oder Beobachtungen in Thüringen (Hildeb
randt & Semm ler 1976). Nach Schmidt (1986) erfolgten von 1946 bis 1980 198
Beobachtungen mit insgesamt 631 Exemplaren (vgl. auch Abb. 3). Ab den 1980er Jahren nehmen
die Beobachtungen in ganz Thüringen stetig zu.
Erste Brutversuche, die aber erfolglos blieben, fanden ab 1987 und in den folgenden Jahren
statt. 2005 brüteten in Ostthüringen 13 und 2006 etwa 25 Brutpaare. Ab 2016 gab es keine Bruten
mehr.
9 Sachsen
Nach Heyde r (1952) ist der Kormoran in Sachsen sehr wahrscheinlich nie Brutvogel gewesen.
Sch lege l (1925) bezeichnet ihn noch als „Irrgast“. In der Vogelwelt Sachsens, die 1998
erschienen ist, wird der Kormoran von Steffe ns et al. als Durchzügler, unregelmäßiger Brutvogel
und Jahresgast bezeichnet. Der erste Nestfund wurde 1977 bekannt. Im Jahr 2007 gab es 222
(LfULG 2009) und 2015 ca. 230 Brutpaare (Deutscher Bundestag 2017).
10 Rheinland-Pfalz
Nach Dolich , Ide lbe rge r & Dietze n (2015) gibt es keine Belege für eine Brut in Rheinland-
Pfalz im 19. Jahrhundert, obwohl der Kormoran schon im Mittelalter auf dem heutigen
Gebiet des Bundeslandes bekannt war.
Der erste rheinland-pfälzische Brutnachweis gelang 1991. Der Bestand lag 2014 bei ca. 320
Brutpaaren. Für das Jahr 2016 werden 420 Brutpaare angegeben (Deutscher Bundestag 2017).
Die Kormoran-Schlafplätze im Winter weisen seit Mitte der 1990er Jahre keinen Anstieg des
Gesamtbestandes von ca. 3.000 Vögeln auf.
11 Saarland
Nach Bos et al. (2005) werden erst seit 1969 Kormorane im Saarland festgestellt. Im Winter
1996 / 97 gab es einen ersten großen Einflug. Seit Ende der 1990er Jahre wurden regelmäßige
Übersommerungen von einzelnen Vögeln oder kleineren Gruppen an verschiedenen Orten des
Saarlandes beobachtet, wobei sie sich meistens an den ausgewählten Winterplätzen aufhalten.
Brutnachweise sind bisher nicht bekannt.
12 Baden-Württemberg
Für Baden-Württemberg ist auch aus früherer Zeit kein Brutvorkommen bekannt (Hölzi nge r
1987).
Die Nachweise von Kormoranen im Winter steigen ab 1975 deutlich über 200 Exemplare und
erreichen im Januar 1985 mit 590 Vögeln ihren vorläufigen Höchstbestand.
Im Atlas der Winterverbreitung der Vögel Baden-Württembergs führen Baue r et al. (1995)
aus: „Status: Brutvogel 1994 bei Karlsruhe und im grenznahen Gebiet der nördlichen Oberrheinebene
in kleiner Zahl; Sommergast in ansteigenden Beständen.“
Für 2015 werden ca. 980 Brutpaare angeführt (Deutscher Bundestag 2017).
13 Bayern
Ob der Kormoran früher in Bayern genistet hat, ist nicht überliefert. 1977 brütete ein Paar erfolgreich
(vgl. Wüst 1982). Ab 1992 / 93 weisen die Kormorane stabile Bestände von ca. 6.127-
7.399 Vögeln auf (Wintermittel von Oktober bis März), bei Maxima von über 9.000 Individuen
Acta ornithoecologica, Bd. 9, H. 1, 2019 67
(vgl. Keller & La nz 2003, Keller & Sch öpf 2005). Nach Bezze l et al. (2005) lag
der gesamte Brutbestand im Jahr 2003 bei ca. 520 Paaren. Im Jahr 2016 betrug er ca. 600 Paare
(Deutscher Bundestag 2017).
Stadtstaaten
Für Hamburg und Umgebung gaben Holzapfe l et al. (1984) für das Jahr 1979 eine Kolonie
mit über 20 Brutpaaren an. Im Jahr 2016 brütet der Kormoran in Hamburg und Berlin mit 400
bzw. 260 BP (Deutscher Bundestag 2017).
Für Deutschland kann aus derzeitiger Sicht eingeschätzt werden, dass die Brutbestände teilweise
rückläufig sind oder stagnieren.
Rutschke (1977) wertete von 1967 bis 1972 die verfügbaren Beobachtungen von Kormoranen
in der DDR aus und schätzte ein: „Winterbeobachtungen (Dezember bis Februar) sind ausgesprochen
selten.“
1997 brüteten in Deutschland ca. 16.450 Brutpaare. Im Jahr 2012 werden für das Gebiet der
Bundesrepublik insgesamt 22.500 Brutpaare in mindestens 150 Kolonien genannt (vgl. Bairlein
et al. 2014). Im Jahr 2015 gab es ca. 24.700 Brutpaare (Deutscher Bundestag 2017). Der
heutige Brutbestand Deutschlands ist höher als je zuvor. Außer in Thüringen, dem Saarland und
Bremen ist der Kormoran in allen Bundesländern Brutvogel. Die meisten Brutvorkommen befinden
sich nach wie vor in den nördlichen Bundesländern Deutschlands.
Seit Jahren sind Kormorane an den unterschiedlichen Gewässern im mitteleuropäischen Binnenland
mehr oder weniger regelmäßige Wintergäste. Keinesfalls ist die Art, auch aus europäischer
Sicht, gefährdet. Für Europa werden für 1992 insgesamt 208.000 Brutpaare angegeben (vgl.
Hegemeije r & Blai r 1997).
3. Zur Situation in Mitteleuropa
Ein Blick in die Schweiz belegt auch dort die positive Bestandsentwicklung bei dieser Art.
Glutz von Blotzheim (1964) schreibt: „Der Kormoran, welcher gegen 1900 noch als seltener
Durchzügler angesprochen wurde und erst seit den Jahren 1923/24 als Wintergast bekannt ist,
überwintert in den letzten Jahren regelmäßig mit mindestens 175 Exemplaren in der Schweiz.“
Erste Brutpaare gab es 2001. Dann erfolgte eine stete Zunahme auf 2.312 BP im Jahr 2017 (vgl.
Müller 2018).
In Österreich brütete der Kormoran nach Aub recht & Böck (1985) bis 1924 an der Lobau,
seit 1945 an der March sowie sporadisch an weiteren zwei Örtlichkeiten. Das Vorkommen in
Niederösterreich erlosch 1971, zwischen 1944 und 1957 gab es auch eine Brutkolonie in Oberösterreich.
Die Autoren führen weiter aus: „Keine andere Wasservogelart hat in den letzten Jahren
so kontinuierlich als Überwinterer zugenommen wie der Kormoran.“ Neben dem Bodensee sind
die Donau und der Inn das zweite große Überwinterungsgebiet. Fließstrecken scheinen von den
Vögeln bevorzugt zu werden.
Im Vorarlberger Rheindelta gab es 2001 die erste erfolgreiche Brut, dann kam es zu einer
raschen Bestandszunahme auf bis zu 200 Nester im Jahr 2008 (Kilze r et al. 2011), 2017 gab es
noch 81 Nester (Knaus et al. 2017).
Für Polen geben Tomia Łojc & Sta warcz yk (2003) für das Jahr 1981 ca. 1.470 Brutpaare
an, die vorwiegend im nördlichen Bereich des Landes brüten (vgl. Abb. 7, links). Etwa 14.000 bis
15.000 Brutpaare, die innerhalb der Landesfläche verteilt sind, nennen die Autoren für das Jahr
2000 (vgl. Abb. 7, rechts).
68 Acta ornithoecologica, Bd. 9, H. 1, 2019
Die Küstenländer der Nord- und Ostsee bilden mit den Niederlanden, Dänemark, Polen und
Norddeutschland den Schwerpunkt der Brutvorkommen des Kormorans in Mitteleuropa (vgl.
We rner 1998, Knief 1996).
Für Dänemark werden für 1984 ca. 5.000 und für 2006 etwa 37.000 Brutpaare angegeben. 2012
gab es 27.237 BP und 2016 stieg der Bestand auf 31.682 BP an (vgl. Steffe ns 2017).
Der Brutbestand in den Niederlanden wird 1978 mit rund 5.000 und 1993 mit 20.460 Brutpaaren
angegeben. 1994 lag der Brutbestand bei 14.000 BP und erreichte 1995 16.560 BP (Knief 1996,
Steffe ns 2017).
In Belgien wurden im Jahr 1995 ca. 270 Brutpaare bekannt; in Tschechien und der Slowakei
werden für das gleiche Jahr ca. 280 bzw. 190 Brutpaare genannt (Glutz von Blotzheim
briefl. Angabe 2019).
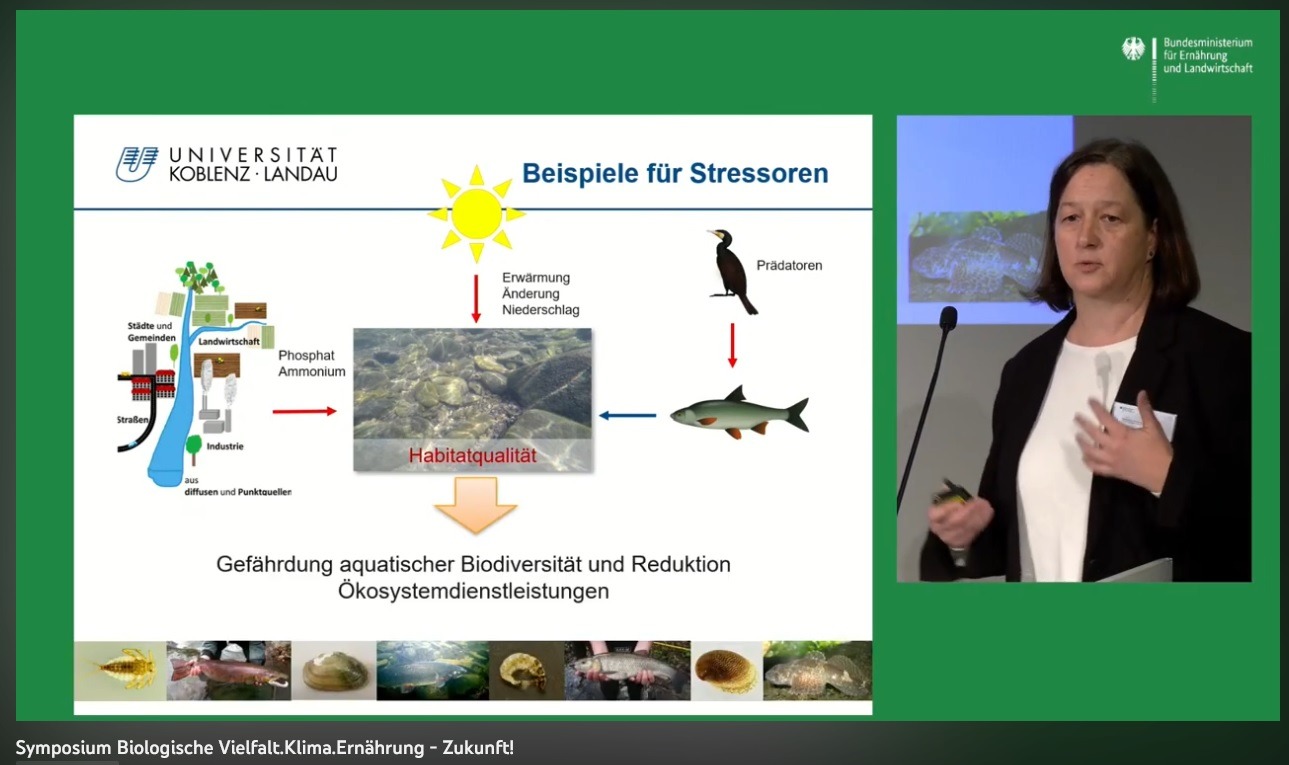
4. Das entstandene Konfliktfeld
Die Zunahme des Kormorans in Europa ab 1980 und auch in Deutschland kann nicht mit Biotopverbesserungsmaßnahmen
erklärt werden (vgl. auch Kap. 2).
Der Artenschutz hat das Ziel und die Verpflichtung, gefährdete Arten zu fördern und durch
Gestaltung der artentsprechenden Biotope zu erhalten. Ökologisch heißt dies bezüglich des
Lebensraumes, dass Generalisten in der Regel nicht gefährdet sind, die Spezialisten hingegen
aber hochgradig.
Das Konfliktfeld bedeutet seit Jahren:
1. Zunahme des Kormoranbestandes außer- und innerhalb des einstigen Brutgebietes im Küstenbereich
von Ost- und Nordsee sowie im unmittelbaren Hinterland
2. Bruten des Kormorans an mehreren Örtlichkeiten im tiefen Binnenland Mitteleuropas und darüber
hinaus
Abb. 7 Verteilung der Brutörtlichkeiten des Kormorans in Polen: links 1981 und rechts im Jahr 2000 (aus Tomia Łojc
& Sta warcz yk 2003).
Acta ornithoecologica, Bd. 9, H. 1, 2019 69
3. Nahezu ganzjährige Anwesenheit der Vögel im Binnenland an den Fließ- und größeren
Standgewässern (Schwerpunkt während der Winterzeit), aber bei saisonal unterschiedlichen
Bestandszahlen
4. Dadurch in jüngster Zeit in den Binnengewässern (Seen, Flüsse, Bäche) verstärkter und ständiger
Prädatorendruck auf bestimmte Fischarten, die sich dort seit Jahrhunderten kaum evolutiv
an den Kormoran anpassen mussten
5. Oft einseitige Auslegung und unzureichendes Wissen über die Problematik der Erhaltung der
Biodiversität in der heutigen Kulturlandschaft
Die Konflikte zwischen Vogel-, Fisch- und Biotopschutz werden von Menschen ausgetragen.
Oberstes Ziel müsste sein, dass sich Natur- und Artenschützer, Jäger, Ornithologen, Förster,
Fischereibiologen, Fischer, Angler, Landwirte, Veterinäre, Zoologen, Ökologen und interessierte
Heimatforscher auf Augenhöhe fachlich austauschen und um Lösungswege ringen.
Öffentlich scheinen die Belange des Vogelschutzes denen des Fischartenschutzes entgegenzustehen.
Wildlebende Fische werden in der Diskussion oft als „herrenlos“ bezeichnet. Dies gilt aber
ebenso auch für die Kormorane. Insofern sind diese Aussagen wirkungslos.
Die Richtlinien der EU sowie die entsprechenden Gesetzlichkeiten des Naturschutzes fordern
die Erhaltung oder Wiederherstellung der „Biologischen Vielfalt“. Dies bedeutet, dass Vogelund
Fischartenschutz gleichwertig sind und die in Rede stehenden Arten ihre volle Berechtigung
erhalten zu werden, haben. Die Wege dazu sind naturbedingt unterschiedlich, denn stets gilt es, die
jeweilige Populationsebene zu betrachten.
Es besteht kein Zweifel darüber, dass seit etwa 1990 eine verstärkte Präsenz von Kormoranen
im deutschen Binnenland an unterschiedlichen Gewässertypen vorhanden ist. Man braucht beispielsweise
nur die Feststellungen von Nauma nn (1817) und Niethamme r (1940), was
die Brutgebiete der Kormorane und die Vögel als Durchzügler betrifft, heranzuziehen. Selbstverständlich
gibt es Verschiebungen in den Vorkommensgebieten.
Es besteht auch kein Zweifel darüber, dass der Kormoran europaweit zu den am besten unter
Kontrolle stehenden Vogelarten zu zählen ist. Der Entwicklungstrend der Vorkommen als Brutvögel,
Durchzügler und Wintergäste ist nicht nur bekannt, sondern es sind auch unzählige legale
und illegale Versuche unternommen worden, die natürliche Bestandsentwicklung zu beeinflussen.
Es sei auch darauf hingewiesen, dass besonders nach längeren Kälteperioden die stehenden
Gewässer, einschließlich der Küstenbereiche, zufrieren. Dann müssen die Vögel auf die offenen
Fließgewässer oder deren Wasserstrecken im Binnenland oder in noch weiter südliche Gebiete
ausweichen.
Somit erhöht sich dort die Kormoranpräsenz erheblich und deutlich für jeden aufmerksamen
Beobachter. Selbst in Großstädten können nunmehr an solchen Flussabschnitten oder in Bereichen
unterhalb von Wehren starke Trupps von 30 Vögeln und mehr von Menschen aus nächster Nähe
beobachtet werden.
Es besteht ebenfalls kein Zweifel darüber, dass Kormorane in Flüssen, Bächen und Standgewässern
des Binnenlandes Einfluss auf die dortigen Fischbestände haben. Hinzu kommen auch die
Belange der Fischzuchtanlagen, einschließlich der Teichwirtschaften.
Natürlich wachsen die Kormoranbestände nicht ins Unendliche, da auch bei dieser Art dichteabhängige
Faktoren wirken. Bisher ist kein Fall bekannt geworden, dass Kormorane eine Fischart in
einem Gewässersystem vernichtet hätten.
Trotzdem darf der Blick auf den Einfluss des Kormorans auf heimische und besonders gefährdete
Fischarten nicht verstellt werden. Auch bei der Fischfauna sind die Biodiversitätsziele zu
erreichen. Oft wird behauptet, dass der Fischbestand eines Gewässers nicht bekannt ist. Hierzu
liegen aus verschiedenen Ländern eingehende seriöse Untersuchungen von Fischereibiologen vor
(vgl. z. B. Keller et al. 1996, Frenz et al. 1997, Honsig -Erlenbu rg & Fried l 1997,
Sch weve rs & Adam 1998, Baars et al. 2001, Blinker 2001).
70 Acta ornithoecologica, Bd. 9, H. 1, 2019
Nach Freyhof (2009) sind zwischen den späten 1980er und mittleren 1990er Jahren nahezu
alle Populationen der Äsche im gesamten Bundesgebiet dramatisch zurückgegangen. Die Art ist
zwar noch weit verbreitet, doch sind die Populationen heute meist klein bis sehr klein.
Offenbar wird in jüngster Zeit zunehmend anerkannt, dass Kormorane wesentlich zum empfindlichen
Rückgang der Äsche und anderer Arten in den Fließgewässern beigetragen haben (vgl.
z. B. Uib lei n et al. 2000, Baars et al. 2001).
5. Gefährdung der Fische
Die unterschiedlichen Gewässerbiotope in Ostdeutschland haben bis 1990 erhebliche Veränderungen
erfahren. Neben dem Gewässerausbau belasteten Abwässer aus Industrie, Landwirtschaft und
kommunalen Bereichen das Fließgewässernetz erheblich. Das hatte unmittelbar negative Auswirkungen
auf die Fisch- und Zoobenthosfauna. Natürlich gab es auch objektbezogene Ausnahmen
und umfangreiche Bemühungen um die Klärung der Abwässer. In Westdeutschland begann
man mit dem Bau von effektiven Kläranlagen aber schon weit vor 1980. Sowohl im Westen wie
auch im Osten Deutschlands wurden Gewässer nach rein technischen Parametern ausgebaut
und unterhalten. Außerdem erfolgte eine Bebauung oft bis weit in die Hochwasserbereiche von
Bächen und Flüssen. In der Regel wurde aber damals nicht gefordert, dass auch Fische sowie
weitere Lebewesen spezialisierte Lebensräume benötigen. Saubere Gewässer waren ein politisches
Ziel. Kläranlagen bringen nach ihrer Inbetriebnahme sichtbare Erfolge. Renaturierungsmaßnahmen,
oft Gegenstand von Auseinandersetzungen, benötigen viel Zeit und sind nicht sofort
sichtbar.
Wenn etwa die wesentlichen Gefährdungsursachen der Fischfauna in den letzten hundert Jahren
in Deutschland zusammengefasst werden, ergibt sich ein differenziertes Bild (vgl. Tab. 2).
Tab. 2 Gefährdungsursachen der Fischfauna in Fließgewässern.
Acta ornithoecologica, Bd. 9, H. 1, 2019 71
Ursachen Auswirkung im Gewässer / Wirkung auf Fische
Gewässerverunreinigungen • Entstehung von Sauerstoffmangel
• Versauerung, Veränderungen des pH-Wertes
• Auslösung von Veralgung und Verschlammung
• R eduzierung oder Ausfall der Fortpflanzung
• Direkte Tötung
Ausbau von Gewässern
sowie Instandhaltungsmaßnahmen
• Beseitigung vorhandener Habitatstrukturen im Gewässer und Verlust
von Reproduktionsstätten und Nahrungsplätzen
• Veränderungen des Bodensedimentes und dadurch Verlust von Laichplätzen
und Habitaten für Jungfische
• Veränderung der Geschiebeführung und somit der Strömungsverhältnisse
• W irkungen auf die Temperaturverhältnisse im Wasser und den Sauerstoffgehalt
Querverbauungen
(Wehre, Sohlabstürze) und
Stauanlagen
• N egativer Einfluss auf den betreffenden Lebensraum der Fischfauna
• Verhinderung von Wanderungen ausgewählter Fischarten und Einengung
des genetischen Austausches
• Verhinderung der Abdrift von Fischen
• Einstellung der Gewässerdynamik mit ihren Konsequenzen für Fische
• Verschiebung des Artenspektrums bei Fischen und Zoobenthos
• Erhöhung des Sauerstoffmangels sowie des pH-Wertes
Zu den häufig vorgebrachten Argumenten für den schlechten Zustand der Fischfauna werden
angesehen:
• Fehlende Durchgängigkeit des Fließgewässers
• Ausbau oder Verbau von Uferbereichen
• Ausgebaute Gewässerstrecken
• Fehlende Unterstände für Fische
• Strukturarme Gewässersohle
• Diffuser Eintrag von Pestiziden
• Diffuse bis deutliche Nährstoffeinträge
• Funktionslose Gewässerrandstreifen
• Tiefenerosion und Strukturdefizite durch künstlich erzeugte Hochwasserereignisse
• Ö rtliche Gewässerversauerung
Diese Hinweise sind berechtigt, benötigen aber viel Zeit bis sie vollständig gelöst und in der
Praxis gegenstandslos geworden sind. Weitere Einzelheiten können in der Wasserrahmenrichtlinie,
deren Ziele aber bisher nicht erreicht wurden, nachgelesen werden.
In den unterschiedlichen Gewässertypen sind beispielsweise auch Mortalitätsfaktoren für
Fische wie Krankheiten, Parasiten, Nahrungsmangel, Temperaturerhöhung, Mikroplastik, Kannibalismus
und Raubfische sowie das Zusammenspiel der einzelnen Faktoren auf die jeweiligen
Fischbestände weitgehend unbekannt. Nicht nur Flussverbauung und Begradigung, sondern auch
die Gewässergüte beeinflussen die Fischfauna. Zu diesen Ursachen des Fischrückgangs kam der
Einfluss des Kormorans in allen Landesteilen ab etwa 1990 noch dazu.
Tab. 2 (Fortsetzung)
72 Acta ornithoecologica, Bd. 9, H. 1, 2019
Ursachen Auswirkung im Gewässer / Wirkung auf Fische
Freizeitaktivitäten • Beunruhigung von Fischen durch Baden
• Einfluss von Bootsfahrern, Kanuten, Floßfahrern auf das Bodensubstrat
bei Niedrigwasser
• Beunruhigung von Fischen bei tragender Eisdecke durch Befahren
oder Schlittschuhlaufen
Landwirtschaft • Beseitigung von Uferrandstreifen
• Eintrag von Feinboden bei fehlenden bzw. zu schmalen Uferstreifen
• Einspülung von Pflanzenschutzmitteln, Dünger und Salzen, wenn
Uferstreifen fehlen
• Verrohrung oder Ausbau von Bächen mit Betonplatten
• W asserentnahme aus Wasserkörpern
• Verlust von Reproduktionsstätten, Nahrungsplätzen und Jungfischhabitaten
• Direkte Schädigung von Fischen
Fischerei • Infolge von Besatzmaßnahmen Einflussnahme auf Fischpopulationen
• Einsatz von nicht heimischen Fischarten
• Selektive Fischentnahme durch Angelfischerei
• Falsche Bewirtschaftung von Angelgewässern und intensive Fischproduktion
Der Kormoran ist ein „Nahrungsopportunist“ und ernährt sich im Wesentlichen von den häufigsten
Fischen eines Gewässers. Der Einfluss dieser Vögel auf die Fischfauna in deutschen
Gewässern muss differenziert betrachtet und bewertet werden. Es gilt zu unterscheiden:
1. Flachgewässer der Nord- und Ostsee mit den entsprechenden Flussmündungen
2. Flüsse im Binnenland
3. Bäche im Binnenland
4. Seen im Binnenland
5. Teiche im Binnenland
6. Künstlich angelegte Staubecken, Stauseen usw.
Die Belange des Fischartenschutzes konzentrieren sich besonders auf die Flüsse (2) und Bäche (3)
der Forellen-, Äschen- und Barbenregionen (vgl. Abb. 8) und auf Seen im Binnenland (4) mit
speziellen Fischarten.
Auch der Besatz beeinflusst die Fischfauna. Einzelne Angelvereine oder Fischereiausübungsberechtigte
führen noch zweifelhafte Besatzmaßnahmen mit Fischen in Gewässern durch (z. B.
mit Regenbogenforellen, fangfähigen Forellen oder Bachsaiblingen), die auch die Fischfauna
erheblich verändern. Natürlich ist auf diesem Gebiet Aufklärungsarbeit bei bestimmten Angelvereinen
im Sinne des Artenschutzes zu leisten.
Die Regenbogenforelle stammt, wie der Bachsaibling, aus Nordamerika. Erstere wurde in
Deutschland ab 1882 und letzterer 1884 in Europa eingeführt. Die heimische Bachforelle ist auch
durch die genannten Besatzmaßnahmen infolge der Konkurrenz durch Regenbogenforellen und
Bachsaiblinge lokal gefährdet.
Abb. 8 Die drei Fischregionen
schematisch mit ihren Bodensubstraten
und Fischarten dargestellt
(aus Görner 2006).
Acta ornithoecologica, Bd. 9, H. 1, 2019 73
Von Oktober bis Mai eines jeden Jahres laichen die betreffenden Fischarten (vgl. Tab. 3). Seit
Anfang der 1990er Jahre gibt es während der Laichzeit dieser Arten eine verstärkte Kormoranpräsenz
an den entsprechenden Fließgewässern im Binnenland. Bis etwa zum Jahr 2000 wurden
Fließgewässer in Ortslagen im Binnenland vom Kormoran gemieden. Nach dieser Zeit können sie
selbst an Gewässern in Großstädten zunehmend von vielen Menschen beim erfolgreichen Fischen
beobachtet werden.
6. Die Wasserrahmenrichtlinie (WRRL)
Die Wasserrahmenrichtlinie (WRRL ) beinhaltet in ihrer Zielstellung eine Reihe von Regelungen,
die auch Aspekte des Naturschutzes berühren. Sie formuliert ein Verschlechterungsverbot mit den
Qualitätskategorien „sehr gut“, „gut“, „mäßig“, „unbefriedigend“ und „schlecht“.
Anhang V der FFH-Richtlinie: Tierart von gemeinschaftlichem Interesse, deren Entnahme aus der Natur und Nutzung
Gegenstand von Verwaltungsmaßnahmen sein können
Tab. 3 Lebensbedingungen einiger typischer Fischarten der Forellen-, Äschen- und Barbenregionen.
74 Acta ornithoecologica, Bd. 9, H. 1, 2019
Fischart Laichzeit Laichorte Laichsubstrat
Verhalten RLD EU
Aal
Anguilla
anguilla
Juli bis
Oktober
Sargassosee
Äsche
Thymallus
thymallus
März bis
Mai
Kieslückensystem
Kiesige bis
feinkiesige
Substrate
Stationäres
Verhalten,
gesellig
lebend
Stark
gefährdet
(2)
Anhang V
der FFHRichtlinie
Bachforelle
Salmo trutta
Oktober bis
Januar
Kieslückensystem
Kiesiger
Untergrund
Stationäres
Verhalten
Ungefährdet
Barbe
Barbus barbus
Mai bis Juli Kiesige
Stellen
Kiesiger
Untergrund
Geselliger
Grundfisch
Vorwarnliste Anhang V
der FFHRichtlinie
Döbel
Squalius
cephalus
April bis
Juni
Steine und
Wasserpflanzen
Steine und
Wasserpflanzen
In der
Jugend
gesellig
Vorwarnliste
Hasel
Leuciscus
leuciscus
März bis
Mai
Sandigkiesige
Stellen
Steine und
Wasserpflanzen
Schwarmfisch
Vorwarnliste
Nase
Chondrostoma
nasus
März bis
April
Kiesige
Stellen
Kiesiger
Untergrund
Gruppenfisch
Vorwarnliste
Quappe
Lota lota
November
bis März
Harter
Untergrund
Steine Ziehen
zum
Laichen in
Gruppen
Vorwarnliste
Zährte
Vimba vimba
Mai bis Juli Flachwasserzonen
Kies oder
Wasserpflanzen
Gesellige
Art
Gefährdet
(3)
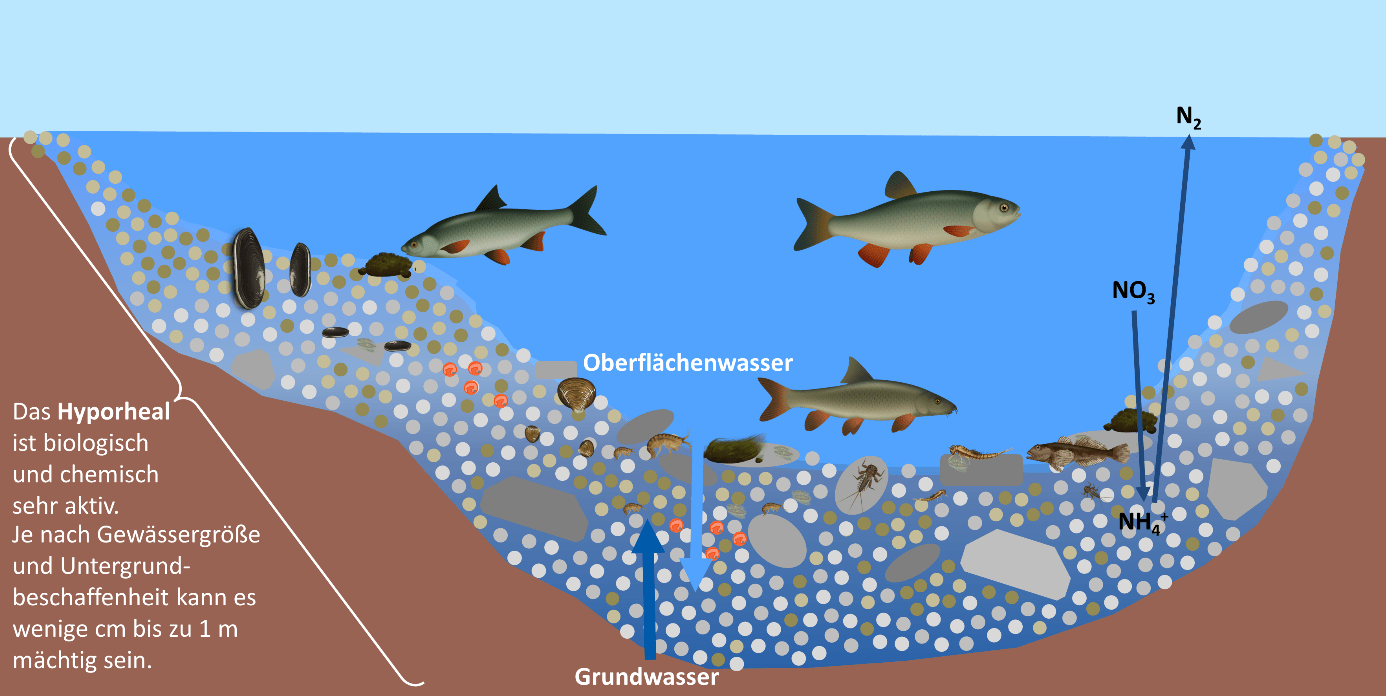
Für die Einstufung der Oberflächengewässer sind selbstverständlich auch biologische Parameter
wie Phytoplankton, Makrophyten, Phytobenthos, Makrozoobenthos und die Fischfauna (siehe
auch Anhang V Nr. 1.2 WRRL ) heranzuziehen.
Der „gute“ Zustand der Gewässer ist an den Fischbeständen festzumachen, die in der Artenzusammensetzung
und Abundanz den typischen Fischgemeinschaften entsprechen und deren Altersstruktur
keine gravierenden Störungen aufweist.
Der „gute Gewässerzustand“ der Oberflächengewässer setzt einen guten ökologischen und
guten chemischen Zustand voraus (vgl. Art. 2 Nr. 18 der WRRL ).
Hier muss beachtet werden, dass das genannte Ziel bei künstlichen oder erheblich veränderten
Wasserkörpern nicht gilt, sondern das eines „guten ökologischen Potentials“ (vgl. Art. 4, Abs. 1a
WRRL ). Solche Gewässerkörper sind beispielsweise Kanäle, Hafenbecken, Talsperren, Baggerseen,
Tagebaurestseen.
Als weitere Faktoren für die Bewertung sind Durchgängigkeit, Morphologie und Hydrologie
heranzuziehen (vgl. Anhang V Nr. 1.2.1 WRRL ). Bei den Fließgewässern müssen Längs- und
Querprofil, Sohlenstruktur, Substrat, Uferbereiche, Laufkrümmung und Strömungsgeschwindigkeit
berücksichtigt werden, wenn es um den ökologischen Zustand geht.
Ebenso sind physikalisch-chemische Parameter zu beachten. Hier gilt es, den Gehalt an Sauerstoff,
Phosphor, Nitrat und Salzen sowie synthetischen und nichtsynthetischen Schadstoffen in
Fließgewässern zu überwachen und die Grenzwerte einzuhalten.
In der WRRL wird zu der Fischfauna hinsichtlich eines „sehr guten Zustands“ ausgeführt:
„Zusammensetzung und Abundanz der Arten entsprechen vollständig oder nahezu vollständig den
Bedingungen bei Abwesenheit störender Einflüsse. Alle typspezifischen störungsempfindlichen
Abb. 9 Ansammlung von Kormoranen
im Stadtgebiet von Gera
im Januar 2006 (Aufn.: H. Ott).
Abb. 10 Vom Kormoran verletzte
Nase. Oft sterben Fische
mit solchen Verletzungen (Aufn.:
K. Schmidt ).
Acta ornithoecologica, Bd. 9, H. 1, 2019 75
Arten sind vorhanden. Die Altersstrukturen der Fischgemeinschaften zeigen kaum Anzeichen
anthropogener Störungen und deuten nicht auf Störungen bei der Fortpflanzung oder Entwicklung
irgendeiner besonderen Art hin.“ Die Aussagen beziehen sich bei allen Zuordnungen der jeweiligen
Zustände auf „anthropogene Störungen“. Bei „mäßigem Zustand“ zeigt „die Altersstruktur
der Fischgemeinschaften […] größere Anzeichen anthropogener Störungen, so dass ein mäßiger
Teil der typspezifischen Arten fehlt oder sehr selten ist.“ Wenn Kormorane in Gewässern durch
Prädation dazu beitragen, dass die gewässerspezifischen Arten (Fische) fehlen oder sehr selten
sind, bleibt dieser Fakt, wie weitere Prädatoren, unberücksichtigt. Würde man der Logik folgen,
wäre das Gewässer als unbefriedigend oder schlecht einzustufen.
Insofern ist es unerheblich, ob anthropogene oder biologische Vorgänge (Prädatoren) auf den
Fischbestand oder besser spezielle Fischarten einwirken. Wäre es nicht ein Gewinn, wenn der
Faktor „Prädation“ reduziert werden könnte? Da es bisher keine effektiven und ökonomisch vertretbaren
Abwehrmaßnahmen gegen Kormorane in den Forellen-, Äschen- und Barbenregionen
des Binnenlandes gibt, bleibt aus derzeitiger Sicht nur eine gewisse Absenkung des Brutbestandes
in einzelnen Ländern Europas. Der Kormoranbestand ist derzeit weder gefährdet, noch besteht
seitens der Jäger jagdliches Interesse an diesem Vogel. Die Abwehrmaßnahmen gegen Kormorane
dürfen aber nicht die einzige praktizierte Maßnahme bleiben.
7. Diskussion
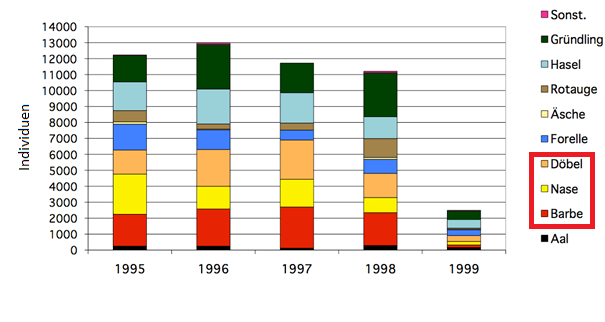
Den Einfluss des Kormorans auf Fischbestände der Forellen-, Äschen- und Barbenregion im Binnenland
haben mehrere Autoren (vgl. z. B. CONR AD et al. 2002, Arge Nister e. V. 2010, KAMMERAD
2015) eindringlich dargestellt. An diesen Fließgewässern ist nunmehr nahezu das ganze
Jahr eine wechselnde Kormoranpräsenz zu beobachten. Bachforellen, Äschen, Barben, Nasen,
Zährten, Hasel, Döbel und gelegentlich Aal sind einem enormen Fraßdruck ausgesetzt, der über
Monate anhält und bei den Äschengewässern einen Bestandsrückgang von 93 bis 99 % der Fischbiomasse
bewirken kann (vgl. BAARS et al. 2001).
Die vorliegenden Beobachtungen belegen, dass bei strengen Wintern eine höhere Kormoranpräsenz
als in milden Wintern an den Flüssen und Bächen des Binnenlandes festzustellen ist. Es
fällt auf, dass die Zeiträume im Sommer oder in wärmeren Wintern, wenn weniger Kormorane
anwesend sind oder festgestellt werden, nicht zur Regenerierung der jeweiligen Fischbestände
ausreichen.
Die Untersuchungen zur Zustandsbestimmung der Gewässer im Rahmen der EU-Wasserrahmenrichtlinie,
die von Fachleuten und nicht von Vertretern der Angelvereine im Auftrag des Staates
durchgeführt werden, bestätigen diese Ergebnisse.
In den oben genannten Gewässerbereichen im Binnenland sind Neubildungen von Brutkolonien
aus den dargelegten Gründen des Fischartenschutzes keinesfalls sinnvoll. Beispielsweise gibt
es in Thüringen derzeit keine Brutplätze des Kormorans. Trotzdem sind einzelne Kormorane an
den genannten Fließgewässern in den Frühjahrs- und Sommermonaten anzutreffen. Hier gilt es
auch, die Verhältnisse in den Schutzgebieten zu prüfen.
In Ostthüringen wurde seit Jahren eine Teichanlage vom NABU übernommen, um den dort auf
Bäumen einer Insel übernachtenden Kormoranen eine „Ablenkfütterung“ anzubieten.
In einem etwa 12 km entfernt befindlichen Fischzuchtbetrieb (Fischteiche) konnte der Fotograf
S. Heidler einen besenderten Kormoran fotografieren (vgl. Abb. 11). Die Auswertung der
Senderdaten ergab, dass der Kormoran täglich zur Nahrungsaufnahme von diesem Schlafplatz
mit unmittelbar vorhandenem Fischteich zum Fischzuchtbetrieb wechselte (S. HEIDLER mündl.
2019). Dieser Fall belegt, dass „Ablenkfütterungen“ für Kormorane eine Wunschvorstellung sind.
Das Vertreiben oder Vergrämen von Kormoranen mit den unterschiedlichen technischen Mitteln
(z. B. Leuchtraketen, Pyrotechnik, Lasergewehre, Gewässerüberspannungen, akustische Signale
u. a.) ist an Fließgewässern nicht zielführend, wie mehrere Beobachtungen aus der Praxis
76 Acta ornithoecologica, Bd. 9, H. 1, 2019
belegen (vgl. auch KELL ER 1996, CONR AD et al. 2002). Diese Methoden der Vergrämung sollten
an Fließgewässern nicht weiter praktiziert werden, da der eindeutige Nachweis ihrer Scheuchwirkungen
auf Kormorane schwer zu erbringen ist und sie für andere Wasservogelarten massive
Störungen bringen können.
Mit den bisherigen Bejagungen des Kormorans, die regelrecht organisiert und von deren Notwendigkeit
Jäger überzeugt werden müssen, sind die Brut- und Rastbestände dieser Vögel nicht
gefährdet.
Kormorane können in der Zeit vom 15. August bis 15. März eines jeden Jahres gejagt werden.
Dies erfordert aber klare staatliche Ausnahmegenehmigungen (siehe Kormoranverordnung). Hier
gilt es, Schutzgebiete, insbesondere internationale und nationale Wasservogelschutzgebiete, von
der Jagd zu verschonen. In den anderen Schutzgebieten mit Fließgewässeranteilen ist zu prüfen,
ob und unter welchen Bedingungen in den Herbst- und Wintermonaten jagdliche Aktivitäten stattfinden
können, ohne dabei die übrigen Wasservögel zu stören.
Oft wird behauptet, aber nicht belegt, dass durch Zuzügler diese Lücken wieder geschlossen
würden.
Das Kormoran-Management muss auf die Bedingungen der regionalen aquatischen Biodiversität
ausgerichtet werden, so dass der Druck von dieser genommen wird. Das ist mit jagdlichen
Maßnahmen sicherlich zu leisten, wenn Jagd als bewusster Arten- und Biodiversitätsschutz praktiziert
wird. Ein lokaler Rückgang von Kormoranen an verschiedenen Gewässern des Binnenlandes
zu den dargelegten Jahreszeiten, bedeutet für die Vögel noch keinerlei Gefährdung.
Unstrittig ist, dass durch die bisherigen Abschüsse lokale Probleme im Artenschutz entschärft
werden konnten.
Natürlich kann ein stärkerer Kormoranbestand seltene oder spezialisierte Fischarten in ihrem
Vorkommen in kurzer Zeit soweit ausdünnen, dass nur noch einzelne, meist aber kleine Fische
auf hunderten von Metern der Gewässerstrecken nachweisbar sind (vgl. z. B. GÖRL ACH &
MÜLL ER 2005, GÖRL ACH & WAGNER 2006, WAGNER et al. 2008).
Die oft geäußerte Meinung, dass die Kormorane durch die Beseitigung von Querbauwerken in
Fließgewässern weniger erfolgreich jagen, ist eine Wunschvorstellung. Beobachtungen belegen,
dass sich ein Trupp von Vögeln teilt und diese sich dann die Fische auf einer Gewässerstrecke
(Bäche und Flüsse) in und gegen die Fließrichtung tauchend und flatternd regelrecht zutreiben.
Anschließend wird der nächste Gewässerabschnitt auf gleiche Weise in Augenschein genommen.
Welche Folgen die Entnahme von Fischen durch die dargelegten Fangaktionen der Kormorane
auf die Alterszusammensetzung der bedrohten Fischpopulationen hat, bleibt unklar. Bei seltenen
Arten kann selbst die Entnahme einzelner Individuen nachhaltige Konsequenzen für den Restbestand
haben.
Es sei mit Nachdruck nochmals betont, dass es landesweit für Vögel ausgewiesene „Vogelschutzgebiete“
gibt. An „Fischschonstrecken“, die gesetzlich festgeschrieben werden und den
Abb. 11 In Polen besenderter Kormoran,
der sich in Thüringen an Teichanlagen
eines Fischzuchtbetriebes bei
Altenburg mehrere Tage aufhielt und
Fische fing (Aufn.: S. heid ler).
Acta ornithoecologica, Bd. 9, H. 1, 2019 77
Schutz der dort vorkommenden Fischarten und des Zoobenthos zum uneingeschränkten Schutzziel
haben, fehlt es weitgehend.
Probleme auszusitzen oder wegzuschauen, ist in der Regel einfacher, aber nicht humaner und
schon keinesfalls hilfreich im Artenschutz.
8. Fazit
Durch den Kormoran sind im Binnenland besonders Fische der Forellen-, Äschen- und Barbenregion
oder weitere autochthone und spezielle Fischarten in Binnenseen gefährdet.
Da eine Reihe von Fischen (vgl. Tab. 3) im Laich-, Ei- und Brutfischstadium an die hochwertigen
Schotterkörper oder ein sauberes Kiessubstrat ökologisch regelrecht gebunden ist, auch wenn
es alters- und artenmäßig Unterschiede gibt, sind diese Fische bereits aufgrund der Veränderungen
im Gewässer (vgl. Tab. 2) bedroht.
Auch in Fließgewässern, wo solche Habitatstrukturen noch weitgehend vorhanden sind und
eine gesicherte Reproduktion der Fische stattfindet, reduzieren Kormorane diese Fischbestände
bis auf wenige Exemplare, wie einschlägige Untersuchungen belegen.
Allein das zeitliche Zusammentreffen von Herbstzug und Überwinterung der Kormorane von
Oktober bis Mitte April an Fließgewässern im Binnenland und beginnender Laichzeit dieser
Fische von Oktober bis Mitte Mai (vgl. auch Tab. 3) bewirkt einen effektiven Eingriff in deren
Populationen. Alle bisherigen Versuche, die fischenden Kormorane von den Fließgewässern fernzuhalten,
blieben erfolglos. Ein effektiver Schutz der Fischfauna in den genannten Fließgewässern
ist ohne Senkung des in das Binnenland einfliegenden Kormoranbestandes nicht möglich.
Es sei mit aller Deutlichkeit darauf verwiesen, dass die derzeit praktizierten Abwehrmaßnahmen
(z. B. Ablenkfütterungen, Einbringen von Totholz in die Gewässer, akustische und optische Vergrämungstechniken,
Entfernen oder Unbrauchbarmachen von Rast- und Schlafplätzen usw.) nicht
zielführend waren.
Eine Bestandsreduktion ist mittelfristig wohl nur erreichbar, wenn Kormorane derzeit im Winterhalbjahr
an den genannten Gewässern erlegt werden. Ferner ist zu prüfen, ob Kormorane im bundesdeutschen
Binnenland, außer denen in den Küstenbereichen und in deren Hinterland, brüten sollten.
Auch der Hinweis auf Vogelprädatoren wie Seeadler (Haliaeetus albicilla), Uhu (Bubo bubo)
und Habicht (Accipiter gentilis) bleibt bei der derzeitigen Bestandsgröße des Kormorans nahezu
wirkungslos. Natürlich kann es auch hier zu weiteren Veränderungen im Nahrungserwerb kommen.
Anders ist das Wirken des Waschbären (Procyon lotor) in Kormorankolonien zu bewerten.
Diese Tiere erklettern die Neststandorte der Vögel auf Bäumen und fressen den Inhalt der Eier
oder die Jungen. Auch die ständige Anwesenheit von Waschbären in den Brutkolonien der Kormorane
kann zu deren Aufgabe führen, wie mehrere Beobachtungen belegen. Ob und welchen
Einfluss dieser oder weitere Prädatoren auf den europäischen Brutbestand des Kormorans haben,
bedarf weiterer Klärung. Hierbei muss natürlich auch bedacht werden, dass der Waschbär für
andere geschützte Arten (z. B. Eulen, Greifvögel, Singvögel, Lurche, Reptilien, Säugetiere, Insekten,
Krebse und Muscheln) ein bestandsbedrohender Prädator ist.
Die deutschen Bundesländer sind für den Vollzug des Artenschutzrechts zuständig und müssen
entscheiden, wann und wie Ausnahmen von den artenschutzrechtlichen Vorschriften erteilt werden
dürfen. Sie können unter den Voraussetzungen des § 45 Abs. 7, Satz 1 des Bundesnaturschutzgesetzes
im Einzelfall Ausnahmen von den Schutzvorschriften für besonders geschützte Tierarten
zulassen. Dies ist der rechtliche Rahmen für Kormoranverordnungen.
Es muss nicht besonders betont werden, dass dem Kormoran auch weiterhin ein gesetzlicher
Schutz zusteht. Dieser muss jedoch flexibel und auf Länderebene, auch im EU-Raum, gestaltbar
sein. Es kann nicht gewartet werden, bis die Revitalisierung der betreffenden Fließgewässer
erreicht ist, um den möglichen Beweis anzutreten, dass dann Kormorane keinen oder nur einen
geringen Einfluss auf die dortige Fischfauna ausüben.
78 Acta ornithoecologica, Bd. 9, H. 1, 2019
In diesem Zusammenhang sei auch auf eine völlig andere Betrachtung der Vogelschutz- und
der FFH-Richtlinie hingewiesen. Die Vogelschutzrichtlinie hat unter anderem zum Inhalt, für die
im Anhang I genannten Arten besondere Schutzmaßnahmen zu ergreifen. Auch die FFH-Richtlinie,
die die Arten in bestimmte Anhänge (Anhang I Lebensraumtypen, Anhang II, IV bzw. V)
einteilt, berücksichtigt eine mögliche Ausbreitung und erhebliche Bestandszunahme bei den Arten
nicht. Was muss im internationalen und nationalen Recht passieren, um aktuellen Situationen bei
diesen Arten gerecht zu werden?
Hierfür gibt es inzwischen zahlreiche Belege aus der Tierwelt, wie beispielsweise Kormoran,
Biber, Fischotter und Wolf, die eine bemerkenswerte Zunahme ihrer Bestände verzeichnen. Diese
Möglichkeit der Bestandszunahme sehen die juristischen Richtlinien bisher nicht vor. Es gibt
keine speziellen Bestimmungen oder Formulierungen, die nachträgliche Änderungen in den Vorschriften
zulassen, dass eine flexible Umsetzung in der Praxis ermöglicht wird.
Ein weitblickender Artenschutz muss auch rechtlich berücksichtigen, dass möglicherweise
Arten in ihren Beständen infolge von Schutzmaßnahmen stark anwachsen können. Auch für solche
Fälle muss es rechtlich saubere Lösungsansätze geben, damit der Rechtsstaat sofort handeln kann.
Zusammenfassung
Die Bestandszunahme des Kormorans in Europa ab 1990 hat zu Konflikten zwischen den Anliegen des Vogel-, Fisch- und
Biotopschutzes geführt.
In diesem Beitrag liegt der Schwerpunkt auf der Sicherung der Biodiversität. Es geht um die Erhaltung der Kormoranpopulationen
sowie der jeweiligen Fischpopulationen im mitteleuropäischen Binnenland. Hier sind besonders die Fischarten
des Lebensraumes Forellen-, Äschen- und Barbenregion betroffen.
Ausdrücklich sei festzuhalten, dass Abwehrmaßnahmen nur eine Seite der Medaille sind. Die Revitalisierung der
Fließgewässer, der Schutz dieser vor Pestizideintrag und die Beseitigung aller weiteren genannten Ursachen des Fischrückgangs
müssen mit gleicher Intensität vorangetrieben werden. Gerade weil diese Maßnahmen sehr zeitaufwendig sind,
dürfen sie nicht hinausgeschoben werden. Es gilt Einsichten zu erlangen, dass die Artenschutzprobleme so gelöst werden,
dass auch im Anthropozän keine Art eine andere auslöscht.
Dank
Mein besonderer Dank gilt Herrn Prof. Dr. U. Glutz von Blotzheim für seine Unterstützung, hilfreichen Kommentare und
Hinweise zum Manuskript. Herrn S. Heidler danke ich für die Übermittlung der Beobachtungen und die zur Verfügung
gestellten Bilder.
Literatur
Albrecht , J., Schmidt , C., Stratma nn, L., Hofma nn, M., Posse lt, S., We ndler, W., RoSSner, D. &
A. Wachse (2012): Die Wasserrahmenrichtlinie aus Sicht des Naturschutzes. – Naturschutz u. Biol. Vielfalt H. 120,
Bonn-Bad Godesberg.
Ande rse n, O. et al. (2018): Storskarven i nedre deler av Gudbrandsdalslågen – Bestandsstørrelse, diett og jakuttak
2017. – NINA Repport 1542. Norsk institutt for naturforskńing. 34 ff.
Anonymus (1989): Vom Kormoran (Phalacrocorax carbo) in Bayern. – Orn. Mitt. 41, Nr. 8, 178-179.
Arbeitsgruppe Artenschutz Thüringen (2007): Studie Einfluss des Kormorans an der Ulster (Thür.). – Jena (unveröffentl.
Gutachten).
Arge Nister e. V. (2010): Der ökologische Gau für ein Gewässer. – Fischer & Teichwirt 61, 333-334.
Aub recht , G. & F. Böck (1985): Österreichische Gewässer als Winterrastplätze für Wasservögel. – Grüne Reihe
Bundesministeriums Gesundheit und Umweltschutz, Bd. 3. – Wien.
Baa rs, M., Mathes , E., Stei n, H. & U. Stei nhörste r (2001): Die Äsche. – Neue Brehm-Büch. 640. – Hohenwarsleben.
Bae r, J. & M. Konrad (2010): Eintrag von Totholz in Fließgewässern – eine Methode zum Schutz von Fischbeständen
vor der Prädation durch Kormorane? – Vogelwarte 48, 15-20.
BAIRL EIN, F., DIERSCHKE, J., DIERSCHKE, V., SALEWSKI, V., GEITER, O., HÜPPOP, K., KÖPPEN, U. & W.
FIEDLER (2014): Atlas des Vogelzugs. – Wiebelsheim.
BAUER, H.-G. & P. BERTHOL D (1996): Die Brutvögel Mitteleuropas. – Wiesbaden.
Acta ornithoecologica, Bd. 9, H. 1, 2019 79
BAUER, H.-G., BOSCHERT, M. & J. HÖL ZINGER (1995): Die Vögel Baden-Württembergs (Bd. 5). – Stuttgart.
Baue r, K. M. & U. N. Glutz von Blotzheim (1966): Handbuch der Vögel Mitteleuropas (Bd. 1). – Frankfurt / M.
Beike, M. (2012): Die Geschichte der Kormoranfischerei in Europa. – Vogelwelt 133, 1-21.
Berg, R. & J. Bae r (2008): Ergebnisse von Untersuchungen an von Kormoranen genutzten Fischbeständen – Beispiele
aus Baden-Württemberg. – Tagungsband „Kormoran und Fischartenschutz“ 16.2.2008 Stuttgart, Schriftenr. Landesfischereiverband
Baden-Württemberg e.V., H. 3, 27-32.
Berndt , R. & G. Busche (1997): Ornithologischer Jahresbericht für Schleswig-Holstein 1995. – Corax 17, 35-65.
Berndt , R. K. & D. Drenckhah n (1974): Vogelwelt Schleswig-Holsteins. – Kiel.
Bezze l, E. (1994): Kormorane im Binnenland. „Übervermehrung“ oder „einregulierte“ Bestände? – Vogelschutz (Hilpoltstein),
H. 2, 14-17.
BEZZEL, E. (1997): Abschuß der Kormorane? – Kosmos H. 5, 62-64.
Bezze l, E., Geie rsbe rge r, I., Loss ow, G. v. & R. Pfeife r (2005): Brutvögel in Bayern. – Stuttgart.
Bless, R. (1989): Artenschutzproblematik und Erforschungsstand bei Fischen und Rundmäulern. – Schriftenr. Landschaftspfl.
Naturschutz H. 29, 217-221. – Bonn – Bad Godesberg.
BLINKER (2001): Fluß leergefressen. – Jahr-Verlag Hamburg 5, 38.
BOCKWINKEL, G. (2014): Totholz in Bächen und Flüssen. – Natur NRW 39, Nr. 1, 24-27.
Bohl, M. (1997): Erfahrungen mit der Abwehr fischfressender Vögel, insbesondere Kormoranen, in der Wielenbacher
Versuchsanlage. – Fischer & Teichwirt 48, 242-246.
Bokranz, W., Neuma nn, D. & J. Borche rdi ng (1998): Der Kormoran am Niederrhein. Untersuchungen zur
Jagdstrategie und zum Beutefischspektrum. – LÖ BF-Mitt. 2, 41-43.
Borgg reve , B. (1869): Die Vogel-Fauna von Norddeutschland. – Berlin.
Bos, J., Buchheit , M. Austge n, M. & O. Elle (2005): Atlas der Brutvögel des Saarlandes. – Orn. Beobachterring
Saar. – Mandelbachtal.
Brämick, U. (2005): Quantifizierung der Auswirkungen des Kormorans auf die Seen- und Flußfischerei Brandenburgs
am Beispiel des Aals. – Referat im Rahmen der Vortragsveranstaltung der Kormoran-Kommission und des Arbeitsausschusses
für Gewässerschutz „Rückgang von Fischbeständen und Fischerei durch Kormoranfraß“ beim Deutschen
Fischereitag in Bingen, 31. August bis 2. September 2005. – Institut für Binnenfischerei, Potsdam.
BRAUNEIS, W. (1999): Zur Ausbreitung des Kormorans (Phalacrocorax carbo) nach Hessen. – Artenschutzreport 9,
60-63.
Brenner, T. (1989): Review: Fischereiwirtschaftliche Schäden durch Graureiher Ardea cinerea L. und Kormoran
Phalacrocorax carbo L. – Fischökologie 1 (2), 61-71.
Buchheim , A. & J. Bellebaum (1993): Bruten des Kormorans (Phalacrocorax carbo) in Nordrhein-Westfalen.
Entwicklung und Zerstörung der ersten westfälischen Kolonie. – Charadrius 29, 93-97.
Bundesamt für Umweltschutz (ed.; 1987): Kormoran und Fischerei. – Schriftenr. Fischerei 47, 1-56, Bern.
Bunte , K. (2005): Die geflügelte Plage vom Möhnesee. Die Akte Kormoran: Bis zu 1.500 Vögel im Sommer zehren
Fischbestände auf. Plage ernsthaftes Problem für Ruhrverband – Arbeit von 14 Jahren in Gefahr. – Soester Anzeiger
20. August 2005.
Bzoma et al. (2005): Salmonid smolts removal by Great Cormorant (Phalacrocorax carbo) from Katy Rybackie colony
(N Poland) in 2000. – 7th International Conference on Cormorants; 4th Meeting of Wetlands International Cormorant
Research Group; 23-26 November 2005, Villeneuve (VD), Switzerland.
CIMIOTTI, D. V., CIMIOTTI, D. S., OCHMANN , T. & J. KREUZIGER (2013): Ornithologischer Jahresbericht für
Hessen 7 (2005-2010). – Vogel u. Umwelt 20, 83-191.
Clayton, E. D. & J. R. Lovv orn (1997): Predation on fish by cormorants and pelicans in a cold-water river: a field
and modeling study. – Canadian Journal of Fishery and Aquatic Sciences 54, 1480-1493.
Conrad , B., Klinge r, H., Schu lze -Wiehe nbrauck , M. & C. Sta ng (2002): Kormoran und Äsche – ein
Artenschutzproblem. – LÖ BF-Mitt. 27 (1), 46-54.
Dauste r, H. (1987): Schäden an Aalbeständen durch Kormorane in schleswig-holsteinischen Binnengewässern. –
Arbeiten des Deutschen Fischereiverbandes 44, 53-57.
Dersi nske , E. (1991): Erfahrungen mit Kormoranen in der Karpfenteichwirtschaft Blumberger Mühle. – Fischer &
Teichwirt 42, 433.
Dersi nske , E. (2006): Aalertragseinbußen durch Kormoranfraß am Beispiel des Grimnitzsees. – Fischer & Teichwirt 3,
91.
Deufe l, J. (1990a): Beeinträchtigung der Fischerei durch Reiher und Kormorane in Süddeutschland. – In: Scha rf,
W. (Hrsg.): Kormorane und Fischreiher im Spannungsfeld zwischen Fischerei und Naturschutz. – Symposiumsbericht
Linz. Öko-Text 1/90, 33-52.
Deufe l, J. (1990b): Fischereischäden durch Wasservögel. – Fischökol. Aktuell 2, 20-25.
Deutscher Angelfischereiverband (2018): Kormoranmanagement – Deutscher Angelfischereiverband fordert im europäischen
Parlament einen gesamteuropäischen Lösungsansatz. – Fischer & Teichwirt 69, H. 12, 448-449.
Deutscher Bundestag (DB) (2017): Bestandsentwicklung der bundesrepublikanischen Kormoranpopulation und deren
Auswirkung auf die Artenvielfalt in heimischen Gewässern. – Antwort der Bundesregierung, Drucksache 18/11360
vom 2.3.2017. – Berlin.
80 Acta ornithoecologica, Bd. 9, H. 1, 2019
Dietze n, C., Dolich , T., Grunwald, T., Keller, P., Kunz, A., Niehuis , M., Schäf , M., Schm olz, M. &
M. Wag ner (2015): Die Vogelwelt von Rheinland-Pfalz. Bd. 2, Entenvögel bis Storchenvögel. – Beih. 47, Fauna u.
Flora in Rheinland-Pfalz.
Dobrowolski , K. A. & R. Dejt rowski (1997): Conflict between fishermen and cormorants Phalacrocorax carbo
in Poland. – Ekologia Polska 45 (1), 279-283.
Dolich , T., Ide lbe rge r, S. & C. Dietze n (2015): Kormoran Phalacrocorax carbo sinensis (Blumenbach, 1798). –
In: Dietze n, C. et al. (2015): Die Vogelwelt von Rheinland-Pfalz. Bd. 2 Entenvögel bis Storchenvögel (Anseriformes
– Ciconiiformes). – Fauna u. Flora Rheinland-Pfalz, Beih. 47, 408-426. – Landau.
Dornbusch , G. & S. Fische r (2010): Nahrungsuntersuchungen an Kormoranen in Sachsen-Anhalt. – Naturschutz
Land Sachsen-Anhalt 47, 16-25.
Dornbusch , M., Dornbusch , G., Fische r, S. & J. Müller (2012): Nachweise von Atlantik-Kormoranen
Phalacrocorax [carbo] carbo in Sachsen-Anhalt. – Apus 17, 37-42.
DuSSling, U. & R. Berg (2001): Fische in Baden-Württemberg. – Stuttgart, Ministerium für Ernährung und Ländlichen
Raum Baden-Württemberg.
Ebel, G. (2000): Habitatansprüche und Verhaltensmuster der Äsche Thymallus thymallus (Li nnaeus , 1758). Ökologische
Grundlagen für den Schutz einer gefährdeten Fischart. – Halle.
Ebel, G. (2012): Zum Einfluss des Kormorans (Phalacrocorax carbo sinensis) auf Fischbestände in Fließgewässern
Sachsen-Anhalts. – Naturschutz Land Sachs.-Anh. 49, 26-39.
Eberl, R. (1990): Beeinträchtigungen der Fischereigewässer durch Kormorane. – Fischer & Teichwirt 41, 8-9.
Fischereiforschungsstelle des Landes Baden-Württemberg – FFSBW (Hrsg.; 2004): Abschlussbericht über Begleituntersuchungen
im Winter 2003 / 2004 zur Verordnung zur Abwendung erheblicher fischereiwirtschaftlicher Schäden durch
Kormorane sowie zum Schutz der heimischen Tierwelt. – Langenargen.
Frenz, C., Klinge r, H. & H. Schuhmache r (1997): Zwischenbericht zur Situation von Äsche (Thymallus thymallus
L.) und Bachforelle (Salmo trutta L.) in der Lenne (NRW ) – Lebensraum Kormorane und Angelfischerei. – Natur
u. Landschaft 72, H. 9, 401-407.
FRENZ, C., PASTER, M., DARSCHNIK, S., ENGELBERG, K. & H. KLINGER (2004): Fischbestände der Forellenregion
in Nordrhein-Westfalen. – LÖ BF-Mitt. 29, Nr. 4, 29-33.
Freyhof, J. (2009): Rote Liste der im Süßwasser reproduzierenden Neunaugen und Fische (Cyclostomata & Pisces). –
Naturschutz u. Biol. Vielfalt 70 (1), 291-316.
Füllner, G. & V. George (2007): Zum Einfluss des Kormorans (Phalacrocorax carbo sinensis) auf den Fischbestand
der Mulde in Sachsen. – Fischer & Teichwirt 8, 290-294.
Füllner, G., Pfeife r, M. & A. Zarske (2005): Atlas der Fische Sachsens. – Dresden.
GEBHARDT, L. & W. SUNKEL (1954): Die Vögel Hessens. – Frankfurt / M.
GEBLER, R.-J. (2005): Entwicklung naturnaher Bäche und Flüsse. Maßnahmen zur Strukturverbesserung. – Walzbachtal.
Geldhause r, F. (1997): Möglichkeiten zur Verminderung von Kormoranschäden in der Karpfenteichwirtschaft. –
Fischer & Teichwirt 48, 326-330.
Gloe, P. (1996): Kormorane – Phalacrocorax carbo – verzehren Aas. – Orn. Mitt. 46, 321.
Glutz von Blotzheim , U. N. (1964): Die Brutvögel der Schweiz (3. Aufl.). – Aarau.
Goethe , F., Hecke nroth , H. & H. Schuma nn (1978): Die Vögel Niedersachsens und des Landes Bremen. –
Naturschutz Landschaftspfl. Niedersachsen, Sonderreihe B, H. 2.1. – Hannover.
Görlach , J. & R. Müller (2005): Die Bestandssituation der Äsche in Thüringen. – Forschungsbericht im Auftrag des
Thüringer Ministeriums für Landwirtschaft, Naturschutz und Umwelt. – Erfurt (unveröffentl. Gutachten).
Görlach , J. & R. Müller (2008): Die Bestandssituation der Äsche (Thymallus thymallus) in Thüringen. – Artenschutzreport
22, 54-62.
Görlach , J. & F. Wag ner (2006): Überprüfung des winterlichen Kormoraneinflusses auf die Fischbestandssituation
in der Ilm / Thüringen. – Schleusingen / Jena (unveröffentl. Gutachten).
Görlach , J. & F. Wag ner (2008): Überprüfung des winterlichen Kormoraneinflusses auf die Fischbestandssituation
der Ilm / Thüringen. – Artenschutzreport 22, 30-45.
Görner, M. (2006): Der Einfluss des Kormorans (Phalacrocorax carbo) und weiterer piscivorer Vögel auf die Fischfauna
von Fließgewässern in Mitteleuropa. – Artenschutzreport 19, 72-88.
Görner, M. (2014): Warum steht der Kormoran (Phalacrocorax carbo) im Mittelpunkt des öffentlichen Interesses? –
Beitr. Jagd- u. Wildforsch. 39, 317-323.
Graf zu Törring-Jette nbach , H. V., Wu nner, U. & P. Wissmath (1995): Fischbestandsituation vor und nach
den Vergrämungsabschüssen im Winter 1994/95. – Fischer und Teichwirt 46, 335-337.
Grémi llet , D. & D. Schmid (1993): Zum Nahrungsbedarf des Kormorans Phalacrocorax carbo sinensis. – Gutachten,
Ministerium für Natur, Umwelt und Landesentwicklung Schleswig-Holstein.
GRÖ GER, J., KINZELBACH, R., PULS, S. & H. M. WINKLER (2009): Wissenschaftliches Gutachten zur qualitativen
Gefährdungsanalyse (PVA) und Abschätzung einer potenziellen Populationsgröße und Populationsstruktur des Kormorans
in M-V, angelehnt an das Konzept der Mindestgrößen von Populationen (MVP) und die Bewertung des „guten
Zustandes der Population“ des Kormorans (Phalacrocorax carbo sinensis) in M-V mit Ableitung eines theoretischen
Acta ornithoecologica, Bd. 9, H. 1, 2019 81
Managementzieles. – Auftraggeber Ministerium für Landwirtschaft, Umwelt und Verbraucherschutz des Landes M-V,
Auftragnehmer Dr. H. M. Winkler, Institut für Biowissenschaften, Universität Rostock.
Gsch weng, M. & A. von Li ndei ner (2011): Kritische Betrachtungen zum Eintrag von Totholz in Fließgewässer als
eine Methode zum Schutz von Fischbeständen vor der Prädation durch Kormorane. – Ber. Vogelschutz 47/48, 45-56.
Güde , H. & D. Strai le (2016): Bodensee. Ökologie und anthropogene Belastungen eines tiefen Voralpensees. – Limnologie
aktuell Bd. 15, Schweizerbart, Stuttgart.
Gürtler, R. (1990): Fischereischäden durch Kormorane und Fischreiher im Jagd- und Naturschutzrecht. – In: Scha rf,
W. (Hrsg.): Kormorane und Fischreiher im Spannungsfeld zwischen Fischerei und Naturschutz. – Symposiumsbericht
Linz. Öko-Text 1/90, 53-66.
Hagemeije r, W. J. & M. J. Blai r (1997): The EBCC Atlas of European Breeding Birds. – London.
Hasmi , D. (1988): Ökologie und Verhalten des Kormorans Phalacrocorax carbo sinensis im Ismaninger Teichgebiet. –
Anz. orn. Ges. Bayern 27, 1-44.
Hecke nroth , H. & V. Laske (1997): Atlas der Brutvögel Niedersachsens 1981-1985. – Naturschutz Landschaftspfl.
Niedersachsen H. 37, 1-329.
HEINICKE, T. (2005): Zur Situation des Kormorans (Phalacrocorax carbo sinensis) in Mecklenburg-Vorpommern. – Ber.
Vogelschutz 42, 97-122.
Hei nrich , D. (2007): Zum vor- und frühgeschichtlichen sowie neuzeitlichen Vorkommen des Kormorans (Phalacrocorax
carbo) in Schleswig-Holstein und angrenzenden Gebieten. – Schr. Naturwiss. Ver. Schleswig-Holst. 69, 3-14.
Hennicke , C. R. (1912): Handbuch des Vogelschutzes. – Magdeburg.
Herrma nn, C. (2011): Der Kormoran Phalacrocorax carbo sinensis in Mecklenburg und Pommern vom ausgehenden
18. bis zur Mitte des 20. Jahrhunderts. – Vogelwelt 132, 1-16.
Herrma nn, C. (2012): Zur Situation des Kormorans Phalacrocorax carbo sinensis in Mecklenburg-Vorpommern und
im südwestlichen Ostseeraum. – Orn. Mitt. 64, Nr. 1/2, 3-13.
HERZIG, F. & A. BÖHNKE (2007): Fachtagung Kormorane 2006. – BfN-Skripte. – Bonn-Bad Godesberg.
HEYDER, R. (1952): Die Vögel des Landes Sachsen. – Leipzig.
HILDEBRANDT, H. & W. SEMMLER (1976): Ornis Thüringens (Teil 2). – Sonderh. Nr. 3 Thür. Orn. Rundbr.
HOL ZAPFEL, C., HÜPPOP, O. & R. MULSOW (1984): Die Vogelwelt von Hamburg und Umgebung (Bd. II). – Neumünster.
HÖL ZINGER, J. (1987): Die Vögel Baden-Württembergs (Teil 2). – Karlsruhe.
HOMEYER, E. F. v. (1837): Systematische Übersicht der Vögel Pommerns mit Rücksicht auf den allgemeinen Charakter
des Landes, das örtliche und quantitative Vorkommen der Vögel, ihre Lebensart, ihren Zug und ihre Abänderungen,
nebst Beiträgen zur beschreibenden Naturgeschichte. – Anklam.
Honsig -Erlenbu rg, W. & T. Fried l (1997): Einfluß des Kormorans auf die Fischbestände in der mittleren Gail
(Kärnten). – Österreichs Fischerei 50, 113-117.
Hubatsch , H. (1989): Kormorane (Phalacrocorax carbo) am Niederrhein. – Charadrius 25, 13-16.
Hüb ner, T. & D. Putze r (1985): Störungsökologische Untersuchungen rastender Kormorane an niederrheinischen
Kiesseen bei Störungen durch Kiestransport, Segel-, Surf- und Angelsport. – Seevögel 6, 122-126.
HÜRL IMANN , J. & S. STEINER (2015): Trockener Sommer – und dann? – Aqua viva, Zeitschr. Gewässerschutz 57,
H. 5, 12-16.
Iliszk o, L. & K. Kozlowska (2003): Pellet production by Great Cormorants Phalacrocorax carbo sinensis at the
Katy Rybackie colonie, N Poland. – In: Keller et al. (Eds.): Cormorants. – Die Vogelwelt – Beiträge zur Vogelkunde
124 (2003) Supplement, 213-216.
Institut für Binnenfischerei e. V. (Hrsg.; 2011): Fische in Brandenburg. – Potsdam-Sacrow.
JÄHNIG, S., HERING, D. & M. SOMMERHÄUSER (Hrsg.; 2011): Fließgewässer-Renaturierung heute und morgen. –
Limnologie aktuell Bd. 13, Stuttgart.
Janda , J. & P. Musi l (1991): Einfluss der Kormoranbestände auf die Erträge der Teichwirtschaft in Südböhmen. –
Vogelschutz in Österreich 6, 81-85.
Janisch , R. (1980): Ergebnisse der fischereilichen Beweissicherung im Zusammenhang mit der Errichtung des Donaukraftwerkes
Abwinden-Asten. – Naturkundl. Jb. Stadt Linz 26, 31-102.
Jepse n, N. (2005): Vortrag über die aktuellen Ergebnisse von Forschungsarbeiten in Dänemark zum Einfluß des Kormorans
auf Fischbestände, insbesondere bedrohte Fischarten und Wandersalmoniden, im Auftrage des dänischen
Fischereiministeriums und im Rahmen des EU-Projektes „Framework for biodiversity Reconciliation Action Plans“
– AFGN-Tagung in Delmenhorst, 12. März 2005.
Jepse n, N., Rav n, H. D. & S. Pede rse n (2018): Change of foraging behavior of cormorants and the effect on river
fish. – Hydrobiologia 820, 189-199.
JÖBGES, M. (2013): Kormoran. – In: GRÜNEBERG, C., SUDMANN , S. R. sowie WEISS, J., JÖBGES, M., KÖN IG,
H., LASKE, V., SCHMITZ, M. & A. SKIBBE: Die Brutvögel Nordrhein-Westfalens. – NWO & LANUV (Hrsg.),
LWL -Museum für Naturkunde, Münster, 138-139.
JÖBGES, M. M. & P. HErKENR ATH (2017): Zum Vorkommen des Kormorans in Nordrhein-Westfalen. – Natur NRW 42,
Nr. 2, 22-26.
82 Acta ornithoecologica, Bd. 9, H. 1, 2019
Jungwirth , A. (1995): Einfluss des Kormorans auf die Fischerei. – Österr. Fischerei 48, 111-125.
Jungwirth , M. (1984): Die fischereilichen Verhältnisse in Laufstauen alpiner Flüsse, aufgezeigt am Beispiel der österreichischen
Donau. – Österr. Wasserwirtschaft, Jg. 36, H. 5/6, 103-111.
Jungwirth , M. (1991): Sportfischerei und Gewässerökologie. – Vogelschutz in Österr. Nr. 6, 21-30.
Jungwirth , M. & H. Wi nkler (1983): Die Bedeutung der Flussbettstruktur für Fischgemeinschaften. – Österr. Wasserwirtschaft
35, 229-234.
Jungwirth , M., Woschitz , G., Zau ner, G. & A. Jagsch (1995): Einfluß des Kormorans auf die Fischerei. –
Österreichs Fischerei 48, 111-125.
Kai nz, E. (1990): Fischereischäden durch Vögel – insbesondere durch Reiher und Kormorane – in europäischen Binnengewässern.
– In: Scha rf, W. (Hrsg.): Kormorane und Fischreiher im Spannungsfeld zwischen Fischerei und
Naturschutz. – Symposiumsbericht Linz. Öko-Text 1/90, 7-18.
Kai nz, E. (1991): Zur fischereilichen Situation der Gewässer im Bereich Linz. – Öko·L. 13, H. 2, 18-35.
Kai nz, E. (1994): Auswirkungen von Kormoranen auf die Fischbestände von zwei oberösterreichischen Fließgewässern.
– Österr. Fischerei 47, 238-250.
KAMMERAD, B. (2015): Zum Einfluss des Kormorans auf Fischbestände und zur Notwendigkeit von Kormoranabwehr-
und Vergrämungsmaßnahmen für den Erhalt biotoptypischer Fischpopulationen in den Gewässern des Landes
Sachsen-Anhalts. – Beitr. Jagd- u. Wildforsch., Bd. 40, 235-251.
Kamme rad , B., Ellerma nn, S., Mencke , J., Wüstema nn, O. & U. Zuppke (1997): Die Fischfauna von
Sachsen-Anhalt. – Ministerium Raumordnung, Landwirtschaft und Umwelt des Landes Sachsen-Anhalt. – Magdeburg.
Kämmereit, M. (2005): Zur Entwicklung der Fischbestände im Dümmer. – Referat im Rahmen der Vortragsveranstaltung
der Kormoran-Kommission und des Arbeitsausschusses für Gewässerschutz „Rückgang von Fischbeständen und
Fischerei durch Kormoranfraß“ beim Deutschen Fischereitag in Bingen, 31. August bis 2. September 2005. – Niedersächs.
Landesamt Verbraucherschutz und Lebensmittelsicherheit. Hannover.
Keller, T. (1992): Untersuchungen zur Nahrungsökologie von in Bayern überwinternden Kormoranen Phalacrocorax
carbo sinensis. – Diplomarb. Ludwig-Maximilian-Universität Würzburg.
Keller, T. (1993): Untersuchungen zur Nahrungsökologie von in Bayern überwinternden Kormoranen Phalacrocorax
carbo sinensis. – Ornithol. Verh. 25 (2/3), 81-128.
Keller, T. (1995): Food of Cormorants Phalacrocorax carbo sinensis wintering in Bavaria, Southern Germany. – Ardea
83 (1), 185-192.
Keller, T. (1996): Maßnahmen zur Abwehr von Kormoranen – eine Übersicht. – Orn. Anz. 35, 13-23.
Keller, T. (1997): Qualitative und quantitative Untersuchungen zur Nahrungsökologie in Bayern überwinternder Kormorane
(Phalacrocorax carbo sinensis). – Diss. Erlangen-Nürnberg.
Keller, T. (1998): Die Nahrung von Kormoranen (Phalacrocorax carbo sinensis) in Bayern. – J. Orn. 139 (4), 389-400.
Keller, T. & U. La nz (2003): Kormoranmanagement in Bayern. – Vogelwelt 124, SH Kormoran, 339-348.
Keller, T. & T. Vorde rmeie r (1994): Abschlußbericht zum Forschungsvorhaben „Einfluß des Kormorans Phalacrocorax
carbo sinensis auf die Fischbestände ausgewählter bayerischer Gewässer unter Berücksichtigung fischökologischer
und fischereiökonomischer Aspekte.“ – Bayerische Landesanstalt für Fischerei, Starnberg.
Keller, T., Vorde rmeie r, T., von Luk owicz , M. & M. Klei n (1996): Der Einfluß des Kormorans Phalacrocorax
carbo sinensis auf die Fischbestände ausgewählter bayerischer Gewässer unter besonderer Berücksichtigung
fischökologischer und fischereiökonomischer Aspekte. – Orn. Anz. 35, 1-12.
KELL ER, V. & C. MÜLL ER (2015): Bestand und Verbreitung des Kormorans Phalacrocorax carbo in der Schweiz und in
Europa. – Orn. Beobachter 112, H. 4, 259-268.
Kenned y, G. J. A. & J. E. Gree r (1988): Predation by cormorants (Phalacrocorax carbo L.) on the salmonid populations
of the River Bush. – Aquaculture and Fisheries Management 19, 159-170.
Kieckbusch , J. (1993): Beobachtungen zur Nahrungswahl des Kormorans (Ph. c. sinensis) in Umgebung des Naturschutzgebietes
„Oehe-Schleimünde“. – Seevögel 14, 19-22.
Kieckbusch , J. J. & B. Koop (1996a): Brutbestand, Rastverbreitung und Nahrungsökologie des Kormorans (Phalacrocorax
carbo sinensis) in Schleswig-Holstein. – Corax 16, 335-355.
Kieckbusch , J. J. & B. Koop (1996b): Kormoranmonitoring in Schleswig-Holstein. – Vogelwelt 117, 349-354.
Kieckbusch , J. J. & B. Koop (1997): Cormorant Phalacrocorax carbo and fishery in Schleswig-Holstein, Germany.
– Ekol. Polska 45, 287-294.
KILZER, R., WILL I, G. & G. KILZER (2011): Atlas der Brutvögel Vorarlbergs. – Wien.
Kinze lbach , R. (2010): Nomenklatur und Geschichte: Der Kormoran in Mitteleuropa. – Falke 57, 70-78.
Kirby, J. S., Holmes , J. S. & R. M. Sellers (1996): Cormorants Phalacrocorax carbo as fish predators: an appraisal
of their conservation and management in Great Britain. – Biological Conservation 75 (2), 191-199.
Klein, M. (1997): Bericht über die Ergebnisse von Elektrobefischungen in ausgewählten bayerischen Fließgewässern mit
Kormoranpräsenz. – Fischer & Teichwirt 48 (11), 487-490.
Klein, M. (2000): Neubewertung des Einflusses von Kormoranen auf Fischbestände in großen Voralpenseen. – Fischer
& Teichwirt 51 (6), 211-216.
Acta ornithoecologica, Bd. 9, H. 1, 2019 83
KNAUS, P., BRUDERER, D. HAGIST, D., HONOL D, J., MAIER, U., SEGELBACHER, G., ULMER, J. TRÖ SCH, S.
& S. WERN ER (2017): Ornithologischer Rundbrief für das Bodenseegebiet Nr. 226. Bericht über den Sommer und die
Brutzeit 2017. – Ornithologische Arbeitsgemeinschaft Bodensee, Konstanz.
Knief , W. (1994): Zum sogenannten Kormoran-„Problem“. Eine Stellungnahme der Deutschen Vogelschutzwarten zum
Kormoran. – Bestand, Verbreitung, Nahrungsökologie, Managementmaßnahmen. – Natur u. Landschaft 69, 251-258.
Knief , W. (1996): Bestand und Verbreitung des Kormorans in Deutschland. – Vogelwelt 117, 344-348.
KNIEF, W. (1997): Zur Situation des Kormorans (Phalacrocorax carbo sinensis) in Deutschland. – Ber. Vogelschutz 35,
90-105.
Knief , W. & H. Witt (1983): Zur Situation des Kormorans (Phalacrocorax carbo) in Schleswig-Holstein und Vorschläge
für seine künftige Behandlung. – Ber. Dt. Sekt. Rat. f. Vogelschutz 23, 67-69.
KNÖ SCHE, R. (2003): The impact of cormorants on the eel fishery in the River Havel catchment area, Germany. – In:
COW X, I. G. (Edit.): Interactions between fish and bird: implications for management. – Fishing News Books, Blackwell
Science, Oxford, 65-71.
Koch -Tscha nen, C. (1999): Altersabhängige Unterschiede in der räumlichen und zeitlichen Habitatnutzung und im
Fangerfolg von Kormoranen Phalacrocorax carbo in drei nordschweizerischen Überwinterungsgewässern. – Diplomarb.,
Zürich.
Kohl, F. (1996): Kormorane und Fische, Naturschutz und Fischerei. Eine Dokumentation des österr. Kuratoriums für
Fischerei und Gewässerschutz. – Brunn a. G., 1-47.
Kohl, F. (2005): Kormoranschäden an Forellen- und Äschengewässern – Beispiele aus Österreich. –Referat im Rahmen
der Vortragsveranstaltung der Kormoran-Kommission und des Arbeitsausschusses für Gewässerschutz „Rückgang von
Fischbeständen und Fischerei durch Kormoranfraß“ beim Deutschen Fischereitag in Bingen, 31. Aug. bis 2. Sept. 2005.
KRALJ, J. & K. D. FEIGE (2015): Veränderungen im Zugverhalten des Kormorans Phalacrocorax carbo sinensis von den
1930er Jahren bis in die Gegenwart. – Vogelwarte 53, 139-154.
Kreuzige r, J., We rner, M., Haass , N., Haass , C. & H. Siege l (1997): Kormoran Phalacrocorax carbo (Linné
1758). – In: Avifauna von Hessen, Bd. 3, 1-13. – Wiesbaden.
KUBE, J. (2004): 50 Jahre Niederhof – Geschichte einer deutschen Kormorankolonie. – Falke 51, H. 8, 256-262.
LEWIN, W.-C., BOR KMANN , I. & D. RITTERBUSCH (2009): Zur Schädigung von Fischbeständen in ausgewählten
Fließgewässern Sachsen-Anhalts durch Kormorane. – Gutachten im Auftrag des Ministeriums für Landwirtschaft und
Umwelt Sachsen-Anhalts. Institut für Binnenfischerei e. V., Potsdam-Sacrow.
KUHK, R. (1939): Die Vögel Mecklenburgs. – Güstrow.
MALTZAN, A. v. (1848): Verzeichnis der bis jetzt in Mecklenburg beobachteten Vögel. – Arch. Naturgesch. Meckl. 2,
29-48.
Martucci , O., Piet relli, I. & C. Consig lio (1993): Fish otoliths as indicators of the Cormorant Phalacrocorax
carbo diet (Aves, Pelecaniformes). – Bollettino Di Zoologia 60 (4), 393-396.
Mebs , T., Klinge r, H. & T. Hüb ner-Misiak (1993): Der Kormoran in Nordrhein-Westfalen. – LÖ BF-Mitt. 4/93,
44-47.
Mellin, M. & R. Krupa (1997): Diet of Cormorant, based on the analysis of pellets from breeding colonies in NE
Poland. – Suppl. Ric. Biol. Selvaggina 26, 511-515.
Melotti , P., Ronca rati , A., Morde nti , O., Loro, F. & A. Dees (1997): Fish-eating bird predation impact on
intensive farming gilthead seabream (Sparus aurata L.) in the North Adriatic area, Italy. – Suppl. Ric. Biol. Selvaggina.
26, 517-520.
Milde nbe rge r, H. (1982): Die Vögel des Rheinlandes (Bd. I). – Düsseldorf.
Morel, P. (1991): Über den Wandel auf dem Speisezettel der Kormorane. – Jahresber. Ornithol. Ges. Basel 121, 39-40.
Morel, P. (1992): Beuteartenspektrum der Kormorane vom Schlafplatz Kembs in den Wintern 1986/87 bis 1988/89. –
In: Bundesamt für Umwelt, Wald und Landschaft (BUWAL; Hrsg.): Grundlagenberichte zum Thema Kormoran und
Fische. – Schriftenr. Fischerei 50, Bern, 146-157.
Müller, R. (1986): Die Nahrung des Kormorans (Phalacrocorax carbo sinensis) am Bodensee. – Schweiz. Fischereiwissenschaft,
3. Jg., Nr. 1, 1-2.
Müller, W. (1995): Kormoran: auf Seen willkommen, an Flüssen weniger. – Ornis 1995/5, 11-14.
MÜLL ER, C. (2018): Seltene und bemerkenswerte Brutvögel 2017 in der Schweiz. – Ornithol. Beob. 115, 339-352.
Nauma nn, J. A. (1817): Naturgeschichte der Land- und Wasser-Vögel des nördlichen Deutschlands und angrenzender
Länder. Nachtrag 8, Heft 8. – Köthen.
Neh ls, G. & P. Gie napp (1997): Nahrungswahl und Jagdverhalten des Kormorans Phalacrocorax carbo im Wattenmeer.
– Vogelwelt 118, 33-40.
Neubaue r, W., Loose , J. & M. Wi nkler (2017): Die Nahrung des Kormorans Phalacrocorax carbo in der Kolonie
im NSG Krakower Obersee. – Naturschutzarb. Mecklenburg-Vorpommern 60, H. 1/2, 56-63.
Neubau r, F. (1957): Beiträge zur Vogelfauna der ehemaligen Rheinprovinz. – Decheniana 110, 1-278.
NIETHAMMER, G. (1940): Handbuch der Deutschen Vogelkunde (Bd. II). – Leipzig.
Oberle, M. (1997): Die Einhausung von Forellenanlagen zur Abwehr von fischfressenden Vögeln. – Fischer & Teichwirt
48, 330-333.
84 Acta ornithoecologica, Bd. 9, H. 1, 2019
Ornithologischer Beobachterring Saar (Hrsg.; 2005): Atlas der Brutvögel des Saarlandes. – Mandelbachtal.
Österreichisches Kuratorium für Fischerei und Gewässerschutz, ÖKF (Hrsg.; 1996): Kormorane und Fische, Naturschutz
und Fischerei. – Wien.
Ped roll, J. C. & C. Zaugg (1995): Kormoran und Fische. – BUWAL, Schriftenr. Umwelt 242, 94 pp.
Peitzmeie r, J. (1969): Avifauna von Westfalen. – Abh. Landesmus. Naturkunde Münster H. 3, 31. Jg.
Pete rse n, A. (1994): Kampf dem Kormoran. Fischfresser soll aus neuen Kolonien vertrieben werden. – Fischer &
Teichwirt 45, 98.
Pfitz ner, G. (1993): Beitrag der „ARGE OÖ . Wasservögel“ zur Klärung der aktuellen Kormoransituation in Oberösterreich.
– Öko·L. 15, 3-21.
Preuss , D. (2002): Nahrungsökologische Untersuchungen zum Einfluss des Kormorans Phalacrocorax carbo sinensis
auf die Fischerei im Küstenbereich Vorpommerns. – Naturschutzarbeit Mecklenburg-Vorp. 45, 57-67.
Prokop, P. (1980): Der Kormoran (Phalacrocorax carbo sinensis) in Österreich. – Egretta 23, 49-55.
Reichh olf, J. (1990): Verzehren überwinternde Kormorane abnorm hohe Fischmengen? – Mitt. Zool. Ges. Braunau,
Bd. 5, Nr. 9/12, 165-174.
Reichh olf, J. (1993): Der Kormoran (Phalacrocorax carbo) am unteren Inn: Entwicklung der Winterbestände, Ernährung
und die Frage der Verluste für die Fischerei. – Öko·L. 15, 32-37.
Reichh olf, J. (1994): Die Wasservögel am Unteren Inn. Ergebnisse von 25 Jahren Wasservogelzählung: Dynamik der
Durchzugs- und Winterbestände, Trends und Ursachen. – Mitt. Zool. Ges. Braunau 6, 1-92.
Reichh olf-Riehm , H. (1993): Das Verteilungsmuster überwinternder Kormorane (Phalacrocorax carbo) am unteren
Inn im Jänner und Februar 1990. – Öko·L. 15, 28-31.
Rippma nn, U. C. (1990): Probleme um die Situation Kormoran und Fischerei in der Schweiz aus der Sicht des Fischbiologen.
– Öko-Text 1/90, 97-113.
Robi n, K., Graf , R. F. & R. Sch nid rig (2017): Wildtiermanagement. – Bern.
Rüger, A. (1993): Kormoran-Konflikt um Nutzung oder Schutz. – Jb. Naturschutz Landschaftspfl. 48, 79-89.
Ruh lE, C. (1985): Der Einfluß der Kormorane auf die Fischbestände im Linthkanal. – Schweiz. Fischereiwissenschaft
(„Petri-Heil“-Beilage) 2, Nr. 5, 9-10.
Rutschke , E. (1977): Brutverbreitung und Herbst- und Winterbestände einiger Wasservogelarten in der DDR. Zum Zug
des Kormorans (Phalacrocorax carbo L.) durch das Binnenland der DDR. – Potsdamer Forschungen, Wiss. Schriftenr.
Pädagog. Hochschule Karl-Liebknecht, Naturwiss. Reihe B, H. 9, 73-78.
Rutschke , E. (1983): Die Vogelwelt Brandenburgs. – Jena.
Rutschke , E. (1994): „Seeraben“ Schießen oder schützen. – Unsere Jagd 44, 40-42.
Rutschke , E. (1997): Der Kormoran. – Berlin.
Ryslav y, T. (2011): Zur Bestandessituation ausgewählter Vogelarten in Brandenburg – Jahresbericht 2008. – Naturschutz
Landschaftspfl. Brandenburg 20, H. 2, 49-62.
Sächsische Landesstiftung Natur und Umwelt (Hrsg.; 2002): Der Kormoran (Phalacrocorax carbo) im Spannungsfeld
zwischen Naturschutz und Teichbewirtschaftung. – Dresden.
Sächsisches Landesamt für Umwelt, Landwirtschaft und Geologie, LfULG (Hrsg.; 2009): Leitfaden für die teichwirtschaftliche
Nutzung in Europäischen Vogelschutzgebieten in Sachsen. – Dresden.
Schadt , J. (1993): Fische, Neunaugen, Krebse und Muscheln in Oberfranken. – Bayreuth, Bezirk Oberfranken, Fachberatung
für Fischerei (Hrsg.).
Scha low, H. (1919): Beiträge zur Vogelfauna der Mark Brandenburg. – Berlin.
Scha renbe rg, W. & W. Schu ltz (1992): Kormorane Phalacrocorax carbo sinensis und Chlorkohlenwasserstoffe
– Eine Untersuchung zur Ökotoxikologie und Bioindikation. – Seevögel 13, 47-53.
Scha rf, W. (Hrsg.; 1990): Kormorane und Fischreiher im Spannungsfeld zwischen Fischerei und Naturschutz. – Symposiumsbericht,
Linz.
Schieme r, F. (1988): Gefährdete Cypriniden – Indikatoren für die ökologische Intaktheit von Flußsystemen. – Natur u.
Landschaft 63, Nr. 9, 370-373.
Schiffe rli, L. (1984): Kormoran und Fischerei. – Informations-Bull. Schweiz. Vogelwarte Sempach.
SCHLEGEL, R. (1925): Die Vogelwelt des nordwestlichen Sachsenlandes. – Leipzig.
SCHLIEKER, E. (1996): Kormoranschäden in Mecklenburg-Vorpommern, Möglichkeiten und Maßnahmen zur Schadensminderung.
– Fischer & Teichwirt 47, 229-230.
Sch lieke r, E. (2005): Wachsende Probleme mit der Bestandsentwicklung des Kormorans in der Küstenregion des
Landes Mecklenburg-Vorpommern und erkennbare Möglichkeiten zur Entspannung der Situation. – Arbeiten des
Deutschen Fischerei-Verbandes e. V., H. 82, 131-146.
Schma lz, W. & M. Schma lz (2003): Gutachten zum Einfluss des Kormorans (Phalacrocorax carbo sinensis) in der
Saale bei Rudolstadt / Schwarza. – Bauhaus-Universität Weimar (unveröffentl. Gutachten).
Schmidt , K. (1986): Kormoran – Phalacrocorax carbo (L., 1758). – In: Knorre, D. v. u. a. (Hrsg.): Die Vogelwelt
Thüringens. – Jena.
Schmidt , R. (1989): Änderungen im Zugverhalten des Kormorans (Ph. carbo) im Zusammenhang mit seinem Bestandsanstieg.
– Beitr. Vogelk. 35, 199-206.
Acta ornithoecologica, Bd. 9, H. 1, 2019 85
SCHNEIDER, R. (2005): Zwischen den Fronten – Jäger im Konflikt zwischen Kormoranschutz und Fischerei. – Unsere
Jagd 56, H. 9, 4-6.
Sch ratte r, D. & J. Trauttma nnsd orFf (1993): Kormorane Phalacrocorax carbo sinensis an Donau und Enns
in Oberösterreich: Analyse der Speiballen. – Orn. Verh. 25 (2/3), 129-150.
Sch röde r, W., Kohl, F. & S. Hanfland (2007): Kormoran- und Fischbestand. Kritische Analyse und Forderungen
des Landesfischereiverbandes Bayern e. V.– LFV Bayern (Hrsg.), München.
Sch weve rs, U. & B. Adam (1998): Zum Einfluß des Kormorans (Phalacrocorax carbo sinensis) auf die Fischbestände
der Ahr (Rheinland-Pfalz). – Österr. Fischerei 51, 198-210.
Sch weve rs, U. & B. Adam (2003): Zum Einfluß des Kormorans auf die Fischbestände der Unteren Eder (Hessen). –
Fischer & Teichwirt 5/2003, 171-173.
Seiche , K. (2002): Bestand und Nahrungsspektrum des Kormorans (Phalacrocorax carbo sinensis) in den Sächsischen
Teichgebieten als Grundlage für eine Schadensberechnung. – In: Sächsische Landesstiftung Natur und Umwelt (Hrsg.):
Der Kormoran (Phalacrocorax carbo) im Spannungsfeld zwischen Naturschutz und Teichbewirtschaftung. – Dresden,
38-44.
Seiche , K. & A. Wü nsche (1996): Kormoran und Graureiher im Freistaat Sachsen. – Materialien Naturschutz u.
Landschaftspfl. 1/1996. Sächs. Staatsministerium Umwelt u. Landesentwicklung, Dresden.
Sellin, D. (1986): Zur Überwinterung sowie zum Nahrungs- und Schlafplatzverhalten des Kormorans, Phalacrocorax
carbo, am Greifswalder Bodden. – Beitr. Vogelk. 32, 281-294.
Siefke , A. (2009): Zur Bestandsentwicklung der Kormorane (Phalacrocorax carbo) auf und um Rügen. – Acta ornithoecol.
6, H. 4, 183-196.
Skiba, R. (1993): Die Vogelwelt des Niederbergischen Landes. – Jahresber. Naturwiss. Ver. Wuppertal, Beih. 2.
Staub , E. (1992): Einfluß des Kormorans (Ph. carbo) auf Fischbestände und Fangerträge in der Schweiz. – BUWAL
Schriftenr. Fischerei 50, 1-138.
Staub , E., Kräme r, A., Müller, R., Ruh le, C. & J. Wa lte r (1992): Einfluß des Kormorans (Phalacrocorax
carbo) auf Fischbestände und Fangerträge in der Schweiz. – In: Bundesamt für Umwelt, Wald und Landschaft
(BUWAL; Hrsg.): Grundlagenberichte zum Thema Kormoran und Fische. – Schriftenr. Fischerei 50, Bern, 1-139.
STEFFENS, R., SAEMANN , D. & K. GRÖ ßLER (Hrsg.; 1998): Die Vogelwelt Sachsens. – Jena.
Steffe ns, W. (2005): Rückgang von Fischbeständen und Fischerei durch Kormoranfraß. – Öffentliche Vortragsveranstaltung
des DFV auf dem Deutschen Fischereitag 2005 in Bingen. – Fischer & Teichwirt 2005 (11), 414-417.
STEFFENS, W. (2017): Ein richtungsweisendes Positionspapier zur Kormoranproblematik. – Fischer & Teichwirt 68, H.
11, 414-417.
Steffe ns, W. (2018a): Einfluss des Kormorans auf die heimische Fischfauna – Notwendigkeit der Bestandsregulierung.
– Fischer & Teichwirt 69, H. 1, 14-17.
Steffe ns, W. (2018b): Kormoranabschüsse in Bayern. – Fischer & Teichwirt 69, H. 2, 57.
Steffe ns, W. (2018c): Wie geht es mit Kormoran, Biber und Wolf in Deutschland weiter? – Fischer & Teichwirt 69,
H. 7, 253-254.
Steffe ns, W. (2019): Bundesverfassungsgericht: Kormoranverordnung ist rechtens. – Fischer & Teichwirt 70, 55-57.
Stemmer, B. (2012): Flexibilität des Kormorans (Phalacrocorax carbo) beim Nahrungserwerb kann regulierende Maßnahmen
zur Erhaltung von Fischbeständen notwendig machen. – Acta ornithoecol. 7, H. 3, 107-115.
Straka, U. (1991): Verbreitung, sommerliche und winterliche Bestandsentwicklung des Kormorans in Österreich. –
Vogelschutz in Österreich, Nr. 6, 48-63.
Sute r, W. (1989): Bestand und Verbreitung in der Schweiz überwinternder Kormorane Phalacrocorax carbo. – Orn.
Beobachter 86, 25-52.
Sute r, W. (1990): Nahrungsökologie des Kormorans in der Schweiz. – In: Scha rf, W. (Hrsg.): Kormorane und
Fischreiher im Spannungsfeld zwischen Fischerei und Naturschutz. – Symposiumsbericht Linz. Öko-Text 1/90,
67-80.
Sute r, W. (1991a): Beeinträchtigen fischfressende Vogelarten unsere Süßwasserfisch-Bestände? – Vogelschutz in
Österr., Nr. 6, 11-15.
Sute r, W. (1991b): Der Einfluss fischfressender Vogelarten auf Süßwasserfisch-Bestände – eine Übersicht. – J. Orn. 132
(1), 29-45.
Sute r, W. (1991c): Zur Nahrungsökologie des Kormorans in der Schweiz. – Vogelschutz Österreich 6, 75-80.
Sute r, W. (1993): Kormoran und Fische. – Veröff. Naturhist. Mus. Bern, Nr. 1.
TIELKE, S., BREYER, P., HÜTTEMANN , S., JARO CINSKI, W. & D. FEY (2015): Das Äschenhilfsprogramm in Nordrhein-
Westfalen. – Natur NRW 40, Nr. 2, 19-22.
Tomia Łojc , L. & T. Sta warcz yk (2003): Awifauna Polski (Bd. 1). – Wrocław.
Traut Tma nnsd orf, J. & D. Sch ratte r (1993): Beitrag zur Nahrungswahl des Kormorans (Phalacrocorax
carbo) im österreichischen Donausystem. – Öko·L., 21-26.
Trauttma nnsd orf, J., Kollar, H. P. & M. Seite r (1990): Der Kormoran (Phalacrocorax carbo) als Wintergast
an der österreichischen Donau. – Mitt. Zool. Ges. Braunau; Bd. 5, Nr. 9/12, 147-156.
Ubl, C. (2004): Untersuchungen zum Nahrungsspektrum des Kormorans im Bereich des Greifswalder Boddens. – Fischerei
& Fischmarkt in Mecklenburg-Vorp., H. 2, 32-38.
86 Acta ornithoecologica, Bd. 9, H. 1, 2019
Ubl, C. (2018): Europäisches Parlament fordert drastische Reduzierung des Kormoranbestandes in Europa. – Fischer &
Teichwirt 69, H. 8, 297.
Uib lei n, F., Fried l, T. & U. Prochi nig (2002): Fischökologischer und fischereilicher Managementplan für das
Natura 2000-Gebiet Obere Drau. – Klagenfurt.
Uib lei n, F., Jagsch , A., Köss ner, G., Weiss , S., Gollma nn, P. & E. Kai nz (2000): Untersuchungen zu
lokaler Anpassung, Gefährdung und Schutz der Äsche (Thymallus thymallus) in drei Gewässern in Oberösterreich. –
Österreichs Fischerei 53, H. 4, 89-165.
Von Li ndei ner, A. (1997): Fischfressende Vogelarten in Bayern – ein Dauerkonflikt. – Ber. Vogelschutz 35, 107-118.
Von Luk owicz , M. (2002): Kormoranschäden in der Binnenfischerei und Möglichkeiten einer Konfliktlösung. – In:
Sächsische Landesstiftung Natur und Umwelt (Hrsg.): Der Kormoran (Phalacrocorax carbo) im Spannungsfeld zwischen
Naturschutz und Teichbewirtschaftung. – Dresden, 28-37.
VUILL E, T. (2018): Die Äsche im Kanton Bern – wie kann der Bestand gesichert werden? – Aqua viva, Zeitschr. Gewässerschutz
60, H. 1, 26-28.
Wag ner, F., Schma lz, W. & M. Görner (2008): Zum Einfluss des Kormorans (Phalacrocorax carbo) auf den
Fischbestand der Ulster (Thüringen). – Artenschutzreport 22, 1-10.
Waidbache r, H. (1989): Veränderungen der Fischfauna durch Errichtung des Donaukraftwerkes Altenwörth. – Veröff.
österr. MaB-Programms, Bd. 14, 123-161.
WERN ER, M. (1998): Verbreitung und Bestandssituation des Kormorans – Phalacrocorax carbo – in Europa mit einer
Übersicht zur Biologie und Ökologie. – Vogel u. Umwelt 9, 207-215.
WERN ER, M. & K. RICHARZ (1998): Kormoran und Fischerei. – Vogel u. Umwelt 9, H. 5, 263-268.
Winkler, H. M. (2005): Nahrungsuntersuchungen am Kormoran aus Küstengewässern als Grundlage der Bewertung
der Beeinflussung der Fischbestände. – Gutachten im Auftrage des Umweltministeriums des Landes Mecklenburg-Vorpommern,
Staatliches Amt für Umwelt und Natur Rostock (unveröff.).
Winkler, H. M. (2010): Die Nahrung des Kormorans. – Falke 57 (Sonderh.), 21-25.
Wissma nn, P. & K. Limbu rg (1987): Fischereischäden durch Massenansammlungen von Kormoranen. – Fischer &
Teichwirt 38, 310-312.
Wissmath , P. et al. (1991): Verzehren überwinternde Kormorane (Phalacrocorax carbo) abnorm hohe Fischmengen? –
Fischer & Teichwirt 42 (2), 21-23.
Wissmath , P. & U. Wu nner (1996): Kormoranschäden in oberbayerischen Fließgewässern im Winter 1995/96. –
Fischer & Teichwirt 47 (4), 126-129.
Wissmath , P., Wu nner, U. & M. Pav linec (1993): Kormorane in Bayern – Bereicherung der Natur oder eine
Plage? – Fischer & Teichwirt 44 (7), 238-244.
Worthma nn, H. & S. Spratte (1987): Nahrungsuntersuchungen an Kormoranen (Phalacrocorax carbo) – Die
Auswirkungen der Kormorane auf die schleswig-holsteinische Binnenfischerei. – Eine Übersicht. – Gutachten Fischereiamt
Schleswig-Holstein.
Worthma nn, H. & S. Spratte (1990): Nahrungsuntersuchungen an Kormoranen vom Großen Plöner See. – Fischer
& Teichwirt 41 (1), 2-8.
Wu nner, U. & P. Wissmath (2004): Zerstörung der Fischbestände der Amper durch Kormorane. – Fischer & Teichwirt
55 (4), 609-612.
WÜST, W. (Hrsg.; 1982): Avifauna Bavariae, Bd. 1 (2. Aufl.). – München.
Wüstemann, O. (1996): Rote Liste der Fischarten Sachsen-Anhalts – Erkenntniszuwachs, Entwicklungstendenzen und
Vorschläge zum Schutzstatus sowie Maßnahmen des Fischartenschutzes. – Ber. Landesamt Umweltschutz Sachs.-Anh.
H. 21, 46-51.
ZANDER, H. D. F. (1862): Systematische Übersicht der Vögel Mecklenburgs. – Arch. Naturgesch. Meckl. 15, 44-150.
Zau ner, G. (2000): Können Kormorane die fischökologische Funktionsfähigkeit beeinträchtigen? Eine Fallstudie an
der Enns. – Typoskript, Abt. Hydrobiologie, Fischereiwirtschaft und Aquakultur, Universität für Bodenkultur, Wien.
Zij lst ra, M. & M. R. va n Eerde n (1995): Pellet production and the use of otoliths in determining the diet of Cormorants
Phalacrocorax carbo sinensis: trials with captive birds. – Ardea 83 (1), 123-132.
Zimme rma nn, H. (1987): Kormoran – Phalacrocorax carbo (L., 1758). – In: Klafs & Stübs (Hrsg.): Die Vogelwelt
Mecklenburgs (3. Aufl.). – Jena.
Zimme rma nn, H. (1989): Kormoran und Fischerei in der DDR. – Beitr. Vogelkd. 35, 1/4, 193-198.
Martin Görner
Arbeitsgruppe Artenschutz Thüringen e. V.
Thymianweg 25
D – 07745 Jena
E-Mail: ag-artenschutz@freenet.de
Acta ornithoecologica, Bd. 9, H. 1, 2019 87